Your new post is loading...
Your new post is loading...

|
Scooped by
Gilbert C FAURE
May 29, 2015 8:21 AM
|
The TOP 10% information you need!
The scoops deal with published (classical or OPEN) and grey literature (blogs, websites, social networks, press releases) allowing rapid access to recently published relevant information May 29, 2015 you were 26796 visitors, viewing this topic 34.5K times., 4900 scoops May 2025: >8.2K scoops, >98.2 visitors, >177,8 views
|
|
Scooped by
Gilbert C FAURE
March 27, 5:17 AM
|
Mycobacterium tuberculosis (M.tb) is a bacterial pathogen that has evolved in humans, and its interactions with the host are complex and best studied in humans.
Myriad immune pathways are involved in infection control, granuloma formation, and progression to tuberculosis (TB) disease. Inflammatory cells, such as macrophages, neutrophils, conventional and unconventional T cells, B cells, NK cells, and innate lymphoid cells, interact via cytokines, cell-cell communication, and eicosanoid signaling to contain or eliminate infection but can alternatively mediate pathological changes required for pathogen transmission. Clinical manifestations include pulmonary and extrapulmonary TB, as well as post-TB lung disease.
Risk factors for TB progression, in turn, largely relate to immune status and, apart from traditional chemotherapy, interventions primarily target immune mechanisms, highlighting the critical role of immunopathology in TB.
Maintaining a balance between effector mechanisms to achieve protective immunity and avoid detrimental inflammation is central to the immunopathogenesis of TB. Many research gaps remain and deserve prioritization to improve our understanding of human TB immunopathogenesis.
Learn more in #ScienceImmunology on #WorldTBDay: https://scim.ag/4iME6wt
|
|
Scooped by
Gilbert C FAURE
March 21, 6:52 AM
|
What if aging is, in part, an immune system problem?
Yesterday, I shared that the thymus - long thought to be a "forgotten organ" in adults - may actually be a critical driver of immune resilience.
Today, let’s dive into the data from our first Nature paper to see exactly how much it matters and what drives its decline.
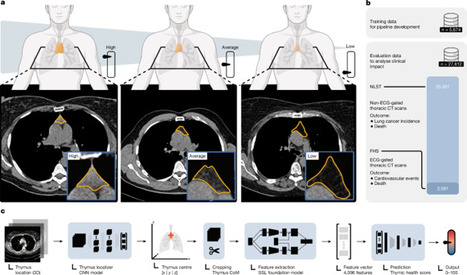
To figure out if thymic health impacts adult longevity, we developed a deep learning system to quantify thymic functionality from routine CT scans. We analyzed over 27,000 asymptomatic adults from two prospective and independent cohorts (the National Lung Screening Trial and the Framingham Heart Study).
The results were striking. We found that the rate of thymic decay is highly individualized - and losing that thymic tissue forecasts a significantly higher risk of disease and death.
👉 Here is what the data showed for individuals with high thymic health compared to those with low thymic health:
- All-cause mortality: An approximately 50% reduction in the risk of death over a 12-year follow-up.
- Cancer: A 36% lower likelihood of developing lung cancer, and nearly a 50% lower likelihood of dying from it. We also saw lower pan-cancer mortality overall.
- Cardiovascular Disease (CVD): Massive risk reductions in CVD mortality ranging from 63% to 92% across the cohorts.
- Other diseases: Mortality from pulmonary diseases was 61% lower, and mortality from endocrine/metabolic diseases was 68% lower.
👉 But here is the also a potential actionable part: Thymic decay isn't just an unavoidable part of aging - it might be tied to how we live.
- Metabolic Health: High-density lipoprotein (HDL) showed a significant positive association with thymic health. Conversely, higher BMI, triglycerides, fasting glucose, and blood pressure were negatively associated with it.
- Lifestyle: Smoking intensity and duration had a strong negative impact on the thymus.
- Inflammation: Participants with chronic systemic inflammation (consistently high CRP levels) or elevated pro-inflammatory proteins (like IL-6 and IL-18) had substantially lower thymic health.
👉 These findings challenge the dogma that the adult thymus is just leftover fatty tissue. If this holds, it positions the thymus as a central regulator of disease susceptibility in adulthood.
Even better, it suggests that actionable lifestyle choices — like smoking reduction, weight loss, and managing inflammation — could potentially preserve or improve our thymic health.
Read the full open-access study here: https://lnkd.in/ePyWdTAM
#Immunology #Longevity #Nature #Aging
|
|
Scooped by
Gilbert C FAURE
March 21, 6:41 AM
|
The thymus, long considered virtually inactive in adulthood—at least that’s how it was presented to me when I was a young medical student—is now confirmed to play a central role in our health and longevity
Numerous studies over the past several years have demonstrated its important role in adults, notably a study published in the NEJM in 2023 that showed the deleterious impact of thymectomy in adults (Kooshesh et al, N Engl J Med 2023 DOI: 10.1056/NEJMoa2302892)
This article published in Nature confirms the importance of the “gland of the soul” (the etymology of “thymus” in Greek) in adults. By analyzing more than 27,000 adults from large cohorts, the authors show that the functional state of the thymus—measured using an innovative artificial intelligence approach applied to CT scans—is strongly associated with the risk of mortality and major diseases.
The results are particularly striking:
Individuals with better “thymic health” have an approximately 50% lower overall risk of death, as well as a significant reduction in the risk of cancer and cardiovascular disease. These associations persist regardless of age, sex, smoking status, or comorbidities, suggesting that the thymus remains a key player in immunity well beyond childhood.
The study also highlights a close link between the thymus, chronic inflammation, and metabolism. Poor thymic health is associated with unfavorable inflammatory profiles, obesity, smoking, and a less healthy lifestyle. Conversely, certain modifiable factors such as physical activity or good metabolic balance appear to preserve its function.
This research reposition the thymus as a major regulator of immune aging. Above all, it opens up significant avenues for further research: better understanding the factors driving its decline, identifying preventive measures, and perhaps one day developing strategies for “immune rejuvenation.” It is worth noting here the likely significant role of obesity, as excess fat in the gland has particularly deleterious effects, and there are interesting studies exploring various strategies involving IL-7, growth hormone, and others.
Ultimately, this study reminds us of the importance of our immune system throughout life and also not to neglect organs considered, without good reason, to be useless. We could also cite interesting studies—even if the impact is less significant than that of the thymus—on the importance, for example, of the tonsils and adenoids or the appendix.
Magali Irla Clémence Granier Daniel Olive
#thymus #health
https://lnkd.in/eEJkY_tE
|
|
Scooped by
Gilbert C FAURE
March 21, 3:50 AM
|
Over the last few months I had the privilege of working together with Victor Appay and Takuya Yamamoto on a review regarding the central importance of CD8⁺ T cells in controlling both acute and chronic viral infections. I am very excited to share that the review is now published in Nature Reviews Immunology.
We discuss qualitative attributes associated with CD8+ T cell efficacy and propose tailored approaches towards interventional strategies
https://lnkd.in/emEYn_Nd
Hope this is helpful both for people working on this field and new comers 😀
#CD8_T_cells_are_cool
#RHIVIERA #ERASE_HIV
Nature Portfolio Institut Pasteur ANRS Maladies infectieuses émergentes The EU2Cure Consortium
|
|
Scooped by
Gilbert C FAURE
March 15, 1:43 AM
|
Neonatal infections remain a leading cause of morbidity and mortality worldwide, reflecting the distinctive immunological state of early life, which p…
|
|
Scooped by
Gilbert C FAURE
March 11, 8:00 AM
|
Access educational materials, eLearning activities, accredited Live webinar sessions with polls and chat on this fast Digital Library and Hybrid Virtual Event Platform powered by MULTILEARNING LMS.
|
|
Scooped by
Gilbert C FAURE
February 27, 3:58 AM
|
|
|
Scooped by
Gilbert C FAURE
February 25, 3:28 AM
|
Interleukin 4 (IL-4) shares a common receptor subunit, IL-4Rα, with IL-13. These two cytokines play an important role in anti-parasitic immune responses. Dysregulated IL-4 /IL-13 expression contributes to Th2-mediated diseases, including asthma and atopic dermatitis.
InvivoGens Recombinant human IL-4 is a high-quality biologically active cytokine, validated using proprietary IL-4/IL-13 reporter cells. This member of the IL-2/γc superfamily is produced in CHO cells to ensure protein glycosylation and bona fide 3D structure (https://lnkd.in/dnemrMTj).
Applications:
- Standard for IL-4 detection and quantification assays
- Screening and release assays for antibodies blocking IL-4 signaling
- Screening and release assays for engineered IL-4
Recombinant human IL-4 can be used together with HEK-Blue™ IL-4/IL-13 cells for the screening of inhibitory molecules, such as Dupilumab, a therapeutic monoclonal antibody targeting the IL-4Rα subunit of the IL-4 receptor (see figures 📊).
#InvivoGen #DrugDiscovery #AsthmaResearch #AtopicDermatisis #Biopharma #Biotech #Lablife
|
|
Scooped by
Gilbert C FAURE
February 21, 4:10 AM
|
Superbe collaboration pour le diagnostic d’un déficit rare en complément chez une jeune fille de 14 ans qui avait été hospitalisée dans le service et qui avait présenté une infection invasive à méningocoque Y
https://lnkd.in/eTZygZmp
💡 toujours faire une évaluation du complément chez les enfants présentant une infection invasive à meningocoque en dosant le C3, C4, CH50 et AP50 et ce, dès le 1er épisode!
💡 recommandation HAS de vaccination par le tétravalent A, C, Y, W à 6 mois et rappel à 12 mois chez tous les enfants
Camille Bougeard Eléonore Eskander @paula viera martins Joan Bitan Julia ROQUIGNY Christèle Kyheng @Veronique Fremeaux-Bacchi, Cécile Gonnin carine el sissy
AP-HP, Assistance Publique - Hôpitaux de Paris
CHU du Kremlin-Bicêtre
#méningocoque #déficitencomplément
#pédiatriegénérale
|
|
Scooped by
Gilbert C FAURE
February 19, 3:18 AM
|
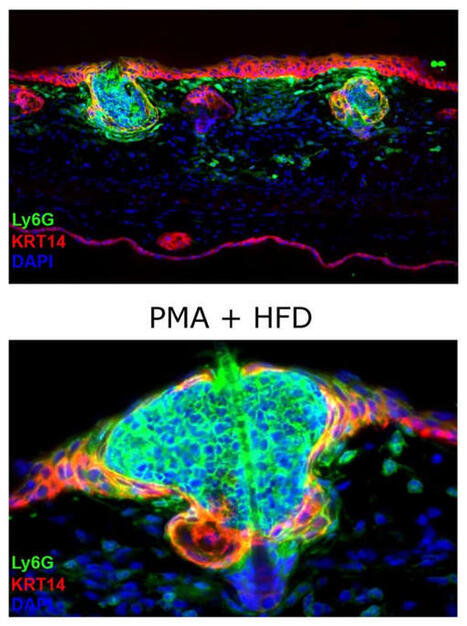
The Trigger: DAMPs and Immunogenic Cell Death
The process begins with tumor cell stress or death (often induced by therapies like radiation or chemotherapy). When these cells die, they release Damage-Associated Molecular Patterns (DAMPs). As shown in the "DAMP signaling" box, this includes:
• Calreticulin: Translocates to the cell surface as an "eat-me" signal.
• HMGB1 & DNA fragments: Passively released from the nucleus/cytoplasm.
• ATP: Actively released to act as a "find-me" signal for immune cells.
These signals activate Dendritic Cells (DCs), which perform cross-presentation—the vital step of taking tumor antigens and presenting them to CD8^+ T cells to "prime" the immune system to recognize the cancer.
2. The cGAS-STING Signaling Pathway
The lower-left portion of the diagram details the intracellular machinery of the cGAS-STING pathway, a primary sensor of cytosolic DNA.
• Detection: In a dying tumor cell or an immune cell that has engulfed tumor DNA, the enzyme cGAS (cyclic GMP-AMP synthase) detects double-stranded DNA in the cytosol.
• Synthesis: cGAS uses ATP and GTP to catalyze the production of cGAMP, a secondary messenger.
• Activation: cGAMP binds to STING (Stimulator of Interferon Genes) located on the endoplasmic reticulum/Golgi apparatus.
• The Signaling Cascade: Activated STING recruits kinases like TBK1 and IKK.
• TBK1 phosphorylates IRF3, which moves into the nucleus.
• IKK leads to the activation of P65 (NF-κB).
• Outcome: These transcription factors trigger the production of Type I Interferons (IFN-I) and pro-inflammatory cytokines such as IL-1\beta, IL-6, and TNF-\alpha.
3. Reprogramming the Microenvironment
The diagram shows that cGAS-STING activation doesn't just kill cells; it reprograms the TME. The "pro-inflammatory TME" at the bottom right is the result of shifting the environment from "cold" (immunosuppressive) to "hot" (immune-active).
• Cellular Reprogramming: The signaling induces the conversion of:
• TAMs (Tumor-Associated Macrophages): Shifting them toward the M1 (anti-tumor) phenotype.
• CAFs (Cancer-Associated Fibroblasts): Modifying their activity to reduce physical barriers to immune entry.
• Tregs & MDSCs: Inhibiting these immunosuppressive cells (Regulatory T cells and Myeloid-Derived Suppressor Cells) that usually help the tumor hide.
|
|
Scooped by
Gilbert C FAURE
February 18, 7:34 AM
|
🔷 Le vieillissement du système immunitaire, est un défi crucial pour la santé des seniors. Des chercheurs se sont penchés sur cette thématique dans un article "Immunofitness in the elderly: The role of vaccination in promoting healthy aging".
Leur objectif, mettre en lumière le rôle capital de la vaccination chez les personnes âgées pour préserver et restaurer l’"immunofitness" sur laquelle nous reviendrons demain.
🔹Le vieillissement induit une transformation profonde du système immunitaire, appelée immunosénescence. Ce processus se caractérise par une diminution progressive de la capacité à générer des réponses immunitaires efficaces face à de nouveaux agents infectieux.
🔹En parallèle, les personnes âgées développent une inflammation chronique de bas grade, qualifiée d’« inflammaging ».
🔹Cette inflammation persistante, alimentée par des cellules immunitaires sénescentes adoptant un profil pro-inflammatoire, ainsi que par des dysfonctionnements au niveau des macrophages et des cellules dendritiques, perturbe l’équilibre immunitaire et favorise l’apparition ou l’aggravation de maladies chroniques.
🔹C'est la combinaison de ces mécanismes qui explique la vulnérabilité accrue des seniors aux infections sévères, notamment respiratoires, leur moindre réponse aux vaccins, ainsi que leur susceptibilité aux complications associées.
🔹Au niveau cellulaire, on a une accumulation de cellules immunitaires qui vieillissent donc, avec des capacités réduites de prolifération, une altération de la présentation antigénique par les cellules dendritiques et une production excessive de cytokines pro-inflammatoires.
🔹Prendre en compte ces altérations immunitaires liées à l’âge est essentiel pour concevoir des interventions ciblées, comme des vaccins adaptés incluant des adjuvants puissants par exemple, et pour développer des stratégies de prévention vaccinale.
Sans oublier des mesures de promotion de la santé comme la nutrition, l'activité physique, afin d’améliorer la qualité de vie de nos seniors !
#santé #prévention #seniors #vaccination #vaccins #santépublique
|
|
Scooped by
Gilbert C FAURE
February 11, 4:15 AM
|
Immunoglobulins Roles In The Immune System
This post explains the roles of all five immunoglobulins in a clear and visual way. IgG provides long term protection, IgA protects mucosal surfaces, IgM signals early infection, IgE mediates allergy and parasite defense, and IgD supports B cell activation. Designed for quick learning, exams, and daily lab reference.
#Immunoglobulins #Antibodies #fblifestyle #IgG #IgA #IgM #IgE #IgD #Immunology #ClinicalImmunology #MLS #MedicalLaboratoryScientist
|
|
|
Scooped by
Gilbert C FAURE
April 1, 3:16 AM
|
Vitiligo is an autoimmune disease of melanocyte destruction, which manifests as progressive, patchy loss of pigmentation in the skin. As one of most common autoimmune diseases, vitiligo inflicts a significant psychosocial burden. Research over the past two decades has revealed the underlying immune mechanisms of vitiligo, with key studies combining detailed analyses of patient tissue samples with mechanistic experiments in mouse models. Vitiligo has emerged as a prototypical CD8+ T cell-mediated autoimmune disease, with cooperation between innate immune cells, dendritic cells, T cells, keratinocytes and fibroblasts driving autoimmune pathology against the uniquely susceptible melanocyte target. The study of vitiligo has also revealed aspects of CD8+ T cell memory and resident memory against self-antigens. This work has drawn from, and contributed to, the study of melanoma immunology. Whereas drugs used for other autoimmune conditions have been largely ineffective in treating vitiligo, a growing base of knowledge recently led to the first successful FDA-approved immune-modulating drugs for vitiligo. This review focuses on the immunology of vitiligo: the mechanisms that drive melanocyte destruction, the biology of aberrant T cell responses against melanocytes and therapeutic means for counteracting this autoimmune condition. This Review from Turk and Huang discusses the immune processes involved in the development of vitiligo, an autoimmune disease in which melanocyte destruction causes loss of skin pigmentation. The authors highlight key studies from the past two decades that have shaped our understanding of vitiligo and led to newly approved immune-modulating drugs for the disease.
|
|
Scooped by
Gilbert C FAURE
March 25, 2:30 PM
|
🛡️ IMMUNE SYSTEM – COMPLETE ANATOMY
🔹 What is Immune System?
The immune system is the body’s defense system that protects against:
Bacteria 🦠
Viruses 🧬
Fungi 🍄
Parasites 🐛
👉 It identifies “self” vs “non-self” and destroys harmful invaders.
🧠 1. MAIN COMPONENTS
🔬 A. Primary Lymphoid Organs
Where immune cells are formed & matured:
✅ Bone Marrow
Produces all blood cells
B-cells mature here
✅ Thymus
T-cells mature here
Active mainly in childhood
🧪 B. Secondary Lymphoid Organs
Where immune response happens:
Lymph nodes
Spleen
Tonsils
Peyer’s patches (intestine)
🧬 2. TYPES OF IMMUNITY
🔹 1. Innate Immunity (Natural)
Present from birth
Fast response
Examples:
Skin barrier
Stomach acid
White blood cells
🔹 2. Adaptive Immunity (Acquired)
Develops over time
Specific + memory-based
Types:
Active (infection/vaccine)
Passive (mother antibodies)
🧪 3. IMMUNE CELLS
🧫 White Blood Cells (WBCs)
🔥 Neutrophils
First responders
Kill bacteria
🧬 Lymphocytes
B-cells → produce antibodies
T-cells → kill infected cells
🧹 Macrophages
Eat pathogens (phagocytosis)
⚔️ Natural Killer (NK) Cells
Kill virus-infected cells
🧴 4. ANTIBODIES (IMMUNOGLOBULINS)
Produced by B-cells
IgG → long-term protection
IgA → mucosal surfaces
IgM → first response
IgE → allergies
IgD → B-cell function
🔥 5. IMMUNE RESPONSE (STEP-BY-STEP)
Pathogen enters body
Recognized as foreign
WBCs activated
Antibodies produced
Pathogen destroyed
Memory cells formed
👉 Next time → faster response ⚡
⚙️ 6. IMPORTANT FUNCTIONS
Protects from infections
Removes dead cells
Fights cancer cells
Creates memory for future protection
🧪 7. COMMON DISORDERS
🔴 Autoimmune Diseases
Body attacks itself
Example: Rheumatoid arthritis
🔴 Immunodeficiency
Weak immune system
Example: HIV/AIDS
🔴 Allergies
Overreaction to harmless substances
🧾 QUICK SUMMARY
FeatureImmune SystemFunctionDefenseCellsWBCsOrgansBone marrow, thymus, lymph nodesTypesInnate & Adaptive
👉 Interested in Medical Coding training?
DM me for complete details. I’ll guide you step-by-step. Feel free to ask your doubts. 9052201814
#MedicalCoding
#HealthcareCareers
#MedicalCoder
#CodingTraining
#CareerGrowth
#OnlineTraining
#JobOpportunity
#WorkFromHome
|
|
Scooped by
Gilbert C FAURE
March 21, 6:48 AM
|
𝐂𝐃𝟕𝟎 𝐢𝐬 𝐞𝐯𝐞𝐫𝐲𝐰𝐡𝐞𝐫𝐞 𝐢𝐧 𝐬𝐨𝐥𝐢𝐝 𝐭𝐮𝐦𝐨𝐫𝐬. 𝐖𝐞 𝐣𝐮𝐬𝐭 𝐰𝐞𝐫𝐞𝐧'𝐭 𝐬𝐞𝐧𝐬𝐢𝐭𝐢𝐯𝐞 𝐞𝐧𝐨𝐮𝐠𝐡 𝐭𝐨 𝐬𝐞𝐞 𝐢𝐭.
In solid tumors, antigen heterogeneity remains a fundamental obstacle. When a target is expressed on only a fraction of tumor cells, CAR T cells eliminate that fraction. The rest survives, proliferates, and drives relapse.
CD70 has long been considered a promising solid tumor target: aberrantly expressed in kidney, ovarian, and pancreatic cancers, while physiologically restricted to activated immune cells. But its heterogeneous expression pattern has limited clinical responses so far.
A new study from Sophie Hanina and al. challenges the way we read that heterogeneity.
𝐓𝐡𝐞 𝐜𝐨𝐫𝐞 𝐡𝐲𝐩𝐨𝐭𝐡𝐞𝐬𝐢𝐬
What if CD70-negative tumor cells aren't truly negative but simply below the detection threshold of conventional methods and conventional CARs?
𝐖𝐡𝐚𝐭 𝐭𝐡𝐞𝐲 𝐟𝐨𝐮𝐧𝐝
🔹 Conventional CD70-CAR T cells selectively killed CD70-high cells and left CD70-low/negative cells untouched, mirroring the modest clinical response rates observed in trials.
🔹 CD70-negative tumor cells were not truly negative. EZH2-mediated H3K27me3 histone trimethylation epigenetically silenced CD70 but incompletely, allowing residual low-level expression to persist across all tumor cells.
🔹 CD70 chromatin accessibility, profiled at single-cell resolution in primary patient tumors, revealed the same epigenetic signature as in PDX models, supporting the existence of pan-cellular, low-level CD70 expression in apparent CD70-negative patient tumors.
𝐄𝐧𝐭𝐞𝐫 𝐭𝐡𝐞 𝐇𝐈𝐓 𝐫𝐞𝐜𝐞𝐩𝐭𝐨𝐫
An HLA-independent T cell (HIT) receptor signals through the CD3 complex, conferring superior sensitivity to low antigen densities.
CD70-HIT T cells, co-expressing CD80 and 4-1BBL for costimulation:
✔ Completely eradicated CD70-heterogeneous tumors across all three cancer models
✔ Showed no greater toxicity than conventional CD70-CAR or CD19-CAR T cells in cytotoxicity assays and in a humanized in vivo model
✔ Spared normal adult tissues, where CD70 chromatin accessibility was largely absent outside activated immune cells
𝐖𝐡𝐚𝐭 𝐭𝐡𝐢𝐬 𝐫𝐞𝐝𝐞𝐟𝐢𝐧𝐞𝐬
Tumor antigen heterogeneity has been framed as a binary problem: cells are either positive or negative. This study reframes it as an epigenetic spectrum and shows that the right receptor design can exploit that spectrum therapeutically.
CD70 may not be a heterogeneous target after all. It may be a pan-cellular target that requires a pan-sensitive receptor.
💬 Has the field been chasing new targets when existing ones just needed a more sensitive receptor? As receptor design becomes more sophisticated, how does this redefine what we manufacture and how we validate it?
📕 Link to the paper in the comments.
|
|
Scooped by
Gilbert C FAURE
March 21, 6:33 AM
|
THYMIC HEALTH IS IN!
For a long time, it has been widely assumed that the thymus plays an important role in childhood, helping build the immune system, and then quietly fades into irrelevance in adulthood. That story is incomplete.
Over the past few years, my colleagues and I set out to revisit this question. The result is something I am both humbled and proud to share: two papers published in Nature, in the same issue, exploring the role of thymic health in adulthood. Beyond the academic milestone, this work represents a shift in how we understand the immune system, aging, and disease risk.
At its core, the message is simple. The thymus does not stop mattering when we grow up. In fact, it may be a key yet overlooked regulator of long-term health.
In our first study, we analyzed more than 27,000 individuals across large, prospective cohorts of generally healthy adults. We found that thymic health varies widely between people of the same age, and that this variation is strongly associated with meaningful outcomes. Individuals with higher thymic health had about a 50% lower risk of death. They were significantly less likely to develop lung cancer and had markedly lower cardiovascular mortality. These are not small effects. Importantly, they persist even after accounting for age, sex, smoking, and comorbidities. Two people can be the same chronological age, yet very different biologically. The thymus appears to be one of the organs shaping that divergence.
In our second study, we asked a different question. If the thymus is central to T cell generation, could its health influence how patients respond to immunotherapy? Most biomarkers today focus on the tumor itself. Important, but incomplete. We wanted to look at the other side of the equation: The Patient. Across a pan-cancer cohort of more than 3,400 patients treated with immune checkpoint inhibitors, we found that thymic health is a strong and consistent predictor of outcomes. Patients with higher thymic health had lower risks of disease progression and death. This was particularly evident in cancers such as lung cancer and melanoma, where immunotherapy is widely used. However, we also observed similar associations in other types of cancer, pointing to a broader, tumor-agnostic signal.
Last but not least, I would like to leave a special word of appreciation and congratulations to my co-first authors, Simon Bernatz, Suraj Pai, and Asbjørn Kjær Attermann, to our supervisors, Hugo Aerts and Nicolai Birkbak, and to all the others involved in this major accomplishment.
#Thymus #HealthyAging #Immunology #PrecisionMedicine #Radiology | 15 comments on LinkedIn
|
|
Scooped by
Gilbert C FAURE
March 20, 11:27 AM
|
Big news: The thymus may be critical for adult health
What if we missed something fundamental about the immune system?
In two back-to-back papers in #Nature we show that the thymus may play a much larger role in health than expected.
Using AI on CT data from 30,000+ people, we found that the health of the thymus varies strongly between people – and is linked to longevity, disease risk, and response to immunotherapy.
So this “forgotten organ” may actually be a key driver of immune resilience throughout life. The implications of this finding could be significant:
• New therapeutic strategies to preserve or restore thymic function
• A new biomarker for aging and disease risk
• Better prediction of immunotherapy outcomes
👉 More to come - we’ll share deeper dives in the coming days
Mass General Brigham, Harvard University, Harvard Medical School, Maastricht University, Mass General Brigham Research, Nature Portfolio, Nature Magazine
Thymic health in adults: https://lnkd.in/ePyWdTAM
Thymic health and immunotherapy outcomes: https://lnkd.in/emcmz5iu | 14 comments on LinkedIn
|
|
Scooped by
Gilbert C FAURE
March 11, 10:33 AM
|
A Content Hub aggregating curated immunology resources, from systemic and mucosal immunity knowledge to clinical and applied immunology (allergy, rheumatology, neurology, biotherapies, laboratory medicine) and societal health concerns (vaccine hesitancy) actualized regularly and offering students, t
|
|
Scooped by
Gilbert C FAURE
February 28, 5:16 AM
|
📚 𝐔𝐧𝐞 𝐦𝐢𝐬𝐞 𝐚̀ 𝐣𝐨𝐮𝐫 𝐦𝐚𝐣𝐞𝐮𝐫𝐞 𝐬𝐮𝐫 𝐥𝐞 𝐒𝐞𝐩𝐬𝐢𝐬 𝐯𝐢𝐞𝐧𝐭 𝐝𝐞 𝐩𝐚𝐫𝐚𝐢̂𝐭𝐫𝐞 𝐝𝐚𝐧𝐬 The Lancet Group (𝐟𝐞́𝐯𝐫𝐢𝐞𝐫 𝟐𝟎𝟐𝟔) !
🔬 Définition & Diagnostic
Le sepsis reste défini comme une dysfonction d'organe menaçant le pronostic vital secondaire à une réponse de l'hôte dérégulée à l'infection. Le diagnostic demeure un défi : les présentations atypiques sont fréquentes, la bactériologie n'est positive que dans 60-70% des cas, et les faux positifs restent nombreux (embolie pulmonaire, lymphome...).
Pas de biomarqueur parfait à ce jour. ⚠️
💉 𝐏𝐫𝐢𝐬𝐞 𝐞𝐧 𝐜𝐡𝐚𝐫𝐠𝐞 : 𝐜𝐞 𝐪𝐮𝐢 𝐜𝐡𝐚𝐧𝐠𝐞 (𝐨𝐮 𝐬𝐞 𝐜𝐨𝐧𝐟𝐢𝐫𝐦𝐞)
🕐 Antibiotiques → Toujours la seule intervention associée de façon constante à une réduction de mortalité.
En choc septique : dans l'heure. En patient non choqué : jusqu'à 5-6h de délai tolérable si le diagnostic est incertain. Dé-escalade dès que possible. Durée : 5-7 jours suffisent dans la majorité des cas.
💧 Remplissage → Jusqu'à 30 mL/kg en phase initiale, mais réévaluation fréquente indispensable. Le bilan hydrique positif persistant est associé à une surmortalité. Penser à la déplétion active dès stabilisation.
💊 Vasopresseurs → Noradrénaline en première ligne, PAM cible 65-70 mmHg. Approche multimodale : Vasopressine en 2ème ligne + Hydrocortisone faible dose (50 mg x4/j) pour améliorer la réponse vasculaire.
🫁 Ventilation → Protection pulmonaire systématique (Vt ~6 mL/kg, Pplat < 30 cmH₂O). Limiter la sédation profonde. Sevrage précoce pour prévenir l'atrophie musculaire.
🍽️ Nutrition → Ne pas nourrir de façon agressive en phase aiguë ! Démarrer l'entérale dans les 3-4 jours. Éviter le suralimentation.
🔮 Perspectives d'avenir
- Biomarqueurs de phénotypage pour personnaliser les thérapies (notamment les corticoïdes)
- Intelligence Artificielle pour la détection précoce
- Focus croissant sur la récupération post-sepsis : 1/6 patients gardent des séquelles sévères, 1/3 décèdent dans l'année
🌍 Ne pas oublier : 85% du fardeau mondial du sepsis concerne les pays à ressources limitées. L'équité d'accès aux soins reste un impératif éthique majeur.
#Réanimation #Sepsis #MédecineIntensive #Lancet #Formation
|
|
Scooped by
Gilbert C FAURE
February 25, 3:56 AM
|
By screening viral proteins’ effects on human cells, researchers in Science discover that immune cells can detect infections not only by recognizing pathogens directly, but also by sensing the damage caused by viral attack.
The findings show that this plant-like immune defense strategy—known as effector-triggered immunity—also operates in mammals.
Learn more: https://scim.ag/4aqbirX
|
|
Scooped by
Gilbert C FAURE
February 24, 9:47 AM
|
The Real Question Is: Who Builds the Immune Intelligence Layer? The race to build the Immunological Digital Twin (IDT) has begun. Pharma wants it to de-risk billion-dollar trials.
|
|
Scooped by
Gilbert C FAURE
February 20, 3:17 AM
|
|
|
Scooped by
Gilbert C FAURE
February 19, 2:57 AM
|
The success of modern vaccines depends not only on antibodies — but on T-helper cell polarization.
Understanding Th1, Th2, and Th17 differentiation is key to designing targeted and durable immunity.
🔬 T-Helper Cell Differentiation: A Central Player in Vaccine Efficacy
The type of T-helper (Th) cell activated during an immune response determines the quality and direction of protection generated after infection or vaccination. Understanding these differentiation pathways is critical for designing next-generation vaccines that induce long-lasting and targeted immunity.
🧬 Naïve CD4+ T cells (Th0) differentiate into distinct subsets depending on the cytokine environment:
🔴 Th1 Cells
Activated in response to intracellular pathogens such as viruses and certain bacteria.
• Driven by IL-12 and IFN-γ
• Characterized by T-bet transcription factor
• Promote macrophage activation and cell-mediated immunity
🟠 Th2 Cells
Activated during extracellular parasitic infections (e.g., helminths).
• Driven by IL-4
• Characterized by GATA3
• Support B-cell activation and antibody production (humoral immunity)
🟣 Th17 Cells
Important in defense against extracellular bacteria and fungi.
• Driven by TGF-β and IL-6
• Characterized by RORγt
• Recruit neutrophils and promote inflammation
🔵 Treg Cells
Maintain immune balance and prevent excessive inflammation.
• Driven by TGF-β and IL-2
• Express Foxp3
• Promote immune regulation and tolerance
💡 Why does this matter for vaccines?
Modern vaccine design aims to direct the immune response toward the most protective Th pathway. For example:
• Viral vaccines often require a strong Th1 response.
• Parasitic infections benefit from Th2-mediated antibody responses.
• Mucosal vaccines may require balanced Th17 activity.
A precise understanding of T-helper cell polarization enables scientists to design smarter adjuvants and achieve durable immunological memory.
#Immunology #Vaccinology #TCells #AdaptiveImmunity #ScientificResearch
|
|
Scooped by
Gilbert C FAURE
February 11, 9:22 AM
|
The Immune System’s “Hidden Organs”: Why Serous Cavities Matter More Than We Thought
This recent review frames the peritoneal, pleural, and pericardial cavities as active immune organs rather than passive fluid spaces. The authors synthesize a decade of work showing that serous cavities are dynamic immune reservoirs, metabolic niches, and communication hubs that shape infection control, fibrosis, cancer spread, and even cardiac and pulmonary disease. For drug developers and translational scientists, this challenges long-standing tissue-centric thinking and opens a largely untapped therapeutic and diagnostic frontier.
Key insights from the review:
◾ Serous cavities host specialized immune ecosystems, including cavity-resident macrophages, B1 cells, innate lymphoid cells, and fat-associated lymphoid clusters (FALCs).
◾ Immune responses in these cavities can rapidly shift from a “fluid phase” to a “solid phase,” with macrophage aggregation, clotting, and tissue tethering acting as first-line containment mechanisms.
◾ Stromal cells (mesothelium, fibroblasts) actively instruct immune identity via retinoic acid, cytokines, and metabolic cues, tightly coupling immunity to tissue repair and fibrosis.
◾ Immune cells traffic bidirectionally between cavities and organs (lung, heart, liver, intestine), blurring classical boundaries between local and systemic immunity.
◾ Sex, age, and developmental origin of immune cells materially influence disease outcomes, with direct implications for translational relevance.
From a therapeutic standpoint, serous cavities represent both a target and a delivery opportunity. These compartments concentrate immune cells, metabolites, and antigens in ways that are distinct from blood or solid tissue. This has implications for immunomodulators, biologics, cell therapies, and even local delivery strategies for inflammatory disease, cancer metastasis, fibrosis, and post-surgical adhesions.
Diagnostics may also evolve: serous fluids could serve as rich, dynamic biomarkers reflecting organ-adjacent immune states more sensitively than peripheral blood.
Most importantly, this work argues for designing drugs and trials that account for cavity-specific immunology rather than assuming uniform tissue behavior.
Diseases like peritonitis, pleural infection, pericarditis, fibrosis, endometriosis, and metastatic cancer all intersect with serous cavity biology. Understanding and therapeutically steering these immune niches could mean earlier intervention, more precise targeting, and fewer off-target effects.
For those in immunology, drug development, or translational medicine, it may be time to start viewing them as actionable immune organs.
Read the full review here: https://lnkd.in/ens2rZhj
#Immunology #DrugDevelopment #TranslationalScience #TissueImmunity #Biotech
|
|
Scooped by
Gilbert C FAURE
February 10, 10:50 AM
|
Within the IL-12 superfamily of heterodimeric cytokines, IL-12 and IL-23, which share a subunit, are among the most pro-inflammatory members. Both are primarily produced by phagocytes and have key roles in activating and regulating T lymphocytes, natural killer cells and innate lymphoid cells. IL-12 predominantly promotes type 1 immune responses, whereas IL-23 is closely associated with type 3 immunity. Their receptors are also heterodimeric and, upon engagement, they trigger ‘cytokine polarization’ (the imprinting of functional identities on immune cells by activating lineage-defining transcription factors), which contributes to inflammation and immunopathology. IL-12 has a key role in various inflammatory conditions and is a potent driver of antitumour immunity, and IL-12 delivery is being explored in several clinical trials in cancer. By contrast, IL-23 is essential for maintaining barrier tissue integrity, yet its dysregulation is a central driver of autoimmune diseases such as psoriasis. Beyond their well-established pro-inflammatory roles, studies of both cytokines have also yielded paradoxical findings. Emerging evidence suggests that both IL-12 and IL-23 can also attenuate immune responses. In this Review, we explore the discovery of IL-12 and IL-23, their canonical pro-inflammatory functions, and recent insights into their immunoregulatory roles in inflammation, cancer and autoimmunity. Here, Becher and colleagues examine the paradoxical roles of IL-12 and IL-23, two IL-12 family cytokines that drive type 1 and type 3 immune responses, respectively. Both promote inflammation by activating T cells, natural cells and innate lymphoid cells through cytokine polarization, yet IL-12 also supports antitumour immunity and IL-23 maintains barrier integrity. This Review highlights emerging evidence that both cytokines can also dampen immune responses, revealing unexpected regulatory roles in cancer, autoimmunity and tissue homeostasis.
|