Your new post is loading...
Your new post is loading...

|
Scooped by
Gilbert C FAURE
December 27, 2013 10:35 AM
|
is the most recent part of Immunology! It appeared less than 40 years ago, while systemic immunity exploded 60 years ago. It is still a minor part of Immunology teaching and research, while the mucosal immune system is at the frontline of encounters with germs, antigens... in other words the environment. major keywords: > 450 posts IgA http://www.scoop.it/t/mucosal-immunity?q=IgA > 125 posts tolerance http://www.scoop.it/t/mucosal-immunity?q=tolerance > 400 posts : microbiome http://www.scoop.it/t/mucosal-immunity?q=microbiome july 2015: almost 2100 scoops, >1700 visitors, >3900 views november 2017 >10K views of >3300 scoops june 2020 >17.6K views, >5.5K visitors, >4.5K scoops may 2024 >22K views, >6.9 visitors, >5.2 scoops
|
|
Scooped by
Gilbert C FAURE
March 27, 5:10 AM
|
World TB Day is a great time to pick up John Green's new book "Everything is Tuberculosis"!
My review of his book in The Lancet Group 👇
https://lnkd.in/gq2eMvPA
|
|
Scooped by
Gilbert C FAURE
March 22, 6:31 AM
|
🧬 Programming lung immunity through mucosal vaccination
A new Science Magazine study shows how intranasal vaccination can induce broad protection against diverse respiratory threats in mice.
Using an intranasal liposomal formulation combining TLR4 and TLR7/8 agonists with antigen, the authors demonstrate durable protection against multiple viral and bacterial respiratory infections, as well as allergic airway inflammation.
Multi-omic profiling of lung tissue reveals several key features of this response:
🔹 Durable tissue-resident T cell immunity
Intranasal vaccination induces persistent antigen-specific CD4⁺ and CD8⁺ tissue-resident memory T cells (TRM) in the lung that remain detectable for months.
🔹 Epigenetic reprogramming of alveolar macrophages
Single-cell transcriptomic and chromatin accessibility analyses reveal sustained transcriptional and epigenomic remodeling of alveolar macrophages, enhancing antigen presentation, phagocytosis, and antiviral responses.
🔹 T cell–innate cell cross-talk via RANKL signaling
Memory T cells imprint macrophage function through RANKL-mediated signaling, establishing a feed-forward circuit between adaptive and innate immunity within lung tissue.
🔹 Rapid spatial immune organization upon infection
Following challenge, vaccinated lungs rapidly form tertiary lymphoid structures, enabling accelerated pathogen-specific T- and B-cell responses.
These findings support the concept of “integrated organ immunity” - a coordinated network of tissue-resident immune and structural cells that can provide broad protection against diverse respiratory threats.
💡 The study also highlights how integrating spatial transcriptomics, single-cell RNA-seq, and chromatin accessibility profiling enables detailed mapping of immune programming directly within lung tissue microenvironments.
📄 Zhang et al., Science (2026)
Mucosal vaccination in mice provides protection from diverse respiratory threats
👉 Read the full study here:
https://lnkd.in/d4uzxQCD
📊 Graphical abstract adapted from the article.
#Immunology #Immunity #SystemsImmunology #Vaccines #SpatialTranscriptomics #SingleCell #Multiomics
|
|
Scooped by
Gilbert C FAURE
March 13, 8:25 AM
|
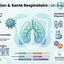
Glycans are essential components of homeostatic networks, acting as fine tuners of immunological responses, and are therefore promising targets for manipulating immune tolerance. Glycans shield the entire gut mucosa surface, contributing to epithelial barrier integrity. Moreover, most microorganisms expose glycoconjugates on their surfaces, making glycans essential molecules in the crosstalk between host immune response and the gut microbiota. The vast amount of biological information encoded by mucosal glycans is deciphered by a variety of glycan-binding proteins that translate glycan recognition into either pro-inflammatory or anti-inflammatory responses. Current evidence from inflammatory bowel disease (IBD) has highlighted the prominent role of glycans in establishing and regulating key cellular and molecular pathways underlying the transition from health to intestinal inflammation, with implications for understanding IBD immunopathogenesis and for IBD prediction and prevention. In this Review, we discuss current advances, emerging challenges and future prospects in exploiting the power of the mucosal glycocalyx and the glycome as master coordinators of the immunoregulatory networks in IBD from the preclinical phase to established diagnosis. We discuss the clinical utility of the glycome as a serological biomarker with diagnostic, prognostic and predictive value, and as a potential new target for preventive intervention strategies in IBD. Glycans are essential components of the gut mucosa that modulate epithelial barrier integrity, host–microbiota interactions and gut immune response. This Review discusses the role of mucosal glycans in gut homeostasis, in intestinal inflammation and their therapeutic potential for inflammatory bowel disease.
|
|
Scooped by
Gilbert C FAURE
March 2, 4:11 AM
|
Un grand merci à Biologiste365 d’avoir permis un débat ouvert et utile autour de “Microbiote intestinal : de la recherche à la clinique”.
Les échanges (et le replay) sont ici : https://lnkd.in/ev3hj_HC
Pour la Société Française de Microbiologie et son GT MicMaC, le message reste constant, déjà porté dans notre tribune (Le Monde, 2023 – https://lnkd.in/ekh2EgXC) : oui à la recherche encadrée sur le microbiote, mais non aux raccourcis, potentiellement dangereux pour les patients.
L’étude portée par MicMaC (Pichon et al., Gut 2025 – https://lnkd.in/eYCweCfW) l’a objectivé : à partir d’un même échantillon de selles standardisé, des offres d’analyse du microbiote “en libre accès” auprès des particuliers produisent à l’heure actuelle des résultats et des interprétations très variables, donc non fiables, et d’aucune utilité. Quand le rendu dépend du laboratoire, ce n’est pas un biomarqueur clinique, et cela ne doit pas guider des décisions de santé.
Ce que nous défendons est simple et pragmatique :
· standardiser (pré-analytique, analytique, bioinformatique)
· valider (recherche clinique, cohortes, réplication, impact clinique)
· encadrer l’interprétation (s’appuyer sur des connaissances médicales et le dialogue clinico-biologique, pas de sur-promesses)
La SFM continuera à pousser une approche exigeante, transparente et centrée sur le patient, pour faire du microbiote un vrai progrès clinique.
Geneviève HÉRY-ARNAUD
#Microbiote #BiologieMédicale #Biomarqueurs #SFM #EBM #Métagénomique #MicMaC
|
|
Scooped by
Gilbert C FAURE
February 21, 4:02 AM
|
|
|
Scooped by
Gilbert C FAURE
February 17, 12:55 PM
|
Harnessing Mucosal Immunity for Protective Vaccines -
A thorough review on mucosal immunity, the type of responses elicited, the unique anatomical and immunological features of the mucosal surfaces of the body, and the challenges associated with the generation of protective immunity via mucosal vaccines.
https://sco.lt/8hqDuy
#vaccines #influenza #Covid19 #RSV #HMPV #HPIV #health #globalhealth #publichealth #medicine #biotechnology #medicine #pharmaceuticals #FDA #CDC #WHO #ECDC
|
|
Scooped by
Gilbert C FAURE
February 15, 1:39 PM
|
What does it mean if we find measles virus in a SEWER? I’m a few weeks into my new role as Vermont’s State Epidemiologist for Infectious Diseases and last week we detected measles virus in wastewater in the state, even though our team hadn’t found any cases of measles since last spring.
Measles virus isn’t usually in wastewater, so finding it there means that someone had measles in the area. They might have been a resident or someone traveling through (and who wouldn’t want to travel through Vermont??).
Fortunately, our epidemiology team has many “streams” of data (get it?). So, a few days later, we were able to identify a case of measles in a person living in the same area. Our epidemiologists connected with the patient to help prevent further spread.
We now know that wastewater is working for us as an early detection system. With rising measles throughout the United States, we will be keeping a close eye on it.
https://lnkd.in/gRQguCwB
|
|
Scooped by
Gilbert C FAURE
February 4, 6:43 AM
|
Each year, vast clouds of dust journey from the Sahara to Europe. But they don't travel alone. They carry a hidden cargo of millions of microbes.
Now, a team from the University of Lisbon, powered by MGI's sequencing tools, is investigating how this invisible migration is reshaping Portuguese agriculture. Their discovery during Storm Célia—a bacterial genus with potential as a powerful bio-fertilizer—turns an environmental phenomenon into a beacon of biotechnological hope.
|
|
Scooped by
Gilbert C FAURE
February 1, 12:53 PM
|
Prior seasonal influenza virus immunity did not impair antibody responses or protection conferred by the intranasal H5N1 vaccine.
|
|
Scooped by
Gilbert C FAURE
January 14, 7:48 AM
|
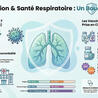
GUT MICROBIOTA IN EARLY CHILDHOOD DEPENDS ON THE MICROBIOTA OF BREAST'S MILK
The establishment of the gut microbiome in early life is critical for healthy infant development.
Mother's milk is crucial for shaping the infant gut microbiome by delivering beneficial bacteria, prebiotics, antibodies, and immune cells, fostering the growth of helpful microbes like Bifidobacterium and reducing pathogens, which is vital for immune development, nutrient absorption, and protection against chronic diseases.
This maternal transfer, via a gut-milk-infant pathway, helps establish a stable, healthy gut ecosystem that supports long-term health.
In an Open Access paper in Nature Communications, the results of an important study on the relationship between intestinal microbiota and breast milk in early childhood.
In this study, the authors quantified the similarity between the maternal milk and the infant gut microbiomes.
They used 507 metagenomic samples collected from 195 mother-infant pairs at one, three, and six months postpartum.
Microbial taxonomic overlap between milk and the infant gut was driven by Bifidobacterium longum, and infant microbiomes dominated by B. longum showed greater temporal stability than those dominated by other species.
They also identified numerous instances of strain sharing between milk and the infant gut, involving both commensal (e.g. B. longum) and pathobiont species (e.g. K. pneumoniae).
Shared strains also included typically oral species such as S. salivarius and V. parvula, suggesting possible transmission from the infant’s oral cavity to the mother’s milk.
At one month, the infant gut microbiome was enriched in biosynthetic pathways, suggesting that early colonisers might be more metabolically independent than those present at six months.
Lastly, they observed significant overlap in antimicrobial resistance gene carriage within mother-infant pairs.
Together, these results suggest that the human milk microbiome has an important role in the assembly, composition, and stability of the infant gut microbiome.
Ferretti, P., Allert, M., Johnson, K.E. et al. Nat Commun 16, 11536 (2025). https://lnkd.in/eD92fRkM
|
|
Scooped by
Gilbert C FAURE
January 12, 8:57 AM
|
Deux études parues dans « Nature Cancer » décrivent comment des niveaux élevés de bactéries infiltrant les tumeurs affaiblissent la réponse immunitaire, favorisant une résistance à l’immunothérapie dans les cancers de la tête et du cou.
|
|
Scooped by
Gilbert C FAURE
January 7, 4:45 AM
|
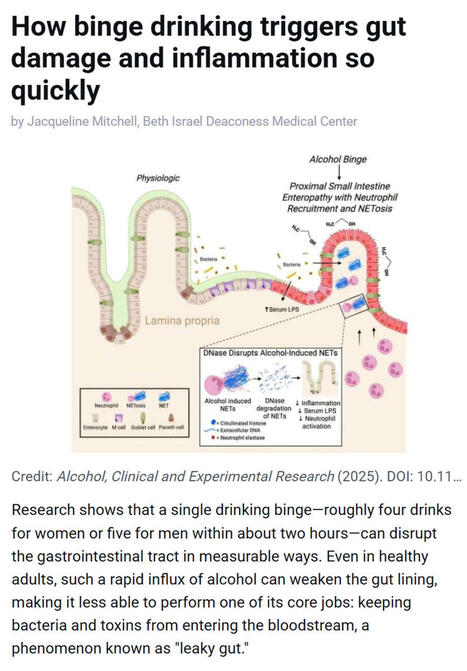
Research shows that even a single episode of binge drinking - about four drinks for women or five for men within two hours - can weaken the gut barrier, allowing #bacteria and #toxins to enter the bloodstream and trigger #inflammation, a process often referred to as “leaky gut.”
▫️ Investigators at Beth Israel Deaconess Medical Center (BIDMC), in work published in Alcohol: Clinical and Experimental Research, found that short bursts of high-dose #alcohol recruit immune cells called #neutrophils to the upper small intestine, where they release damaging structures known as NETs that disrupt the gut lining.
▫️ NETs stands for Neutrophil Extracellular Traps. They are web-like structures made of #DNA, histones, and antimicrobial proteins that are released by neutrophils to trap and kill #microbes. While NETs are part of the body’s innate immune defense, they can also damage surrounding tissues when produced excessively or inappropriately - such as after binge alcohol exposure - by disrupting barriers like the gut lining, promoting inflammation, and allowing bacteria or toxins to leak into the bloodstream.
▫️ The study, led by Scott Minchenberg, MD, PhD, a clinical fellow in #gastroenterology and #hepatology at BIDMC, showed that breaking down these NETs with an enzyme reduced gut damage and bacterial leakage.
▫️ As noted by senior author Gyongyi Szabo MD, PhD, Chief Academic Officer at BIDMC and Beth Israel Lahey Health, these findings highlight an early inflammatory pathway linking binge drinking to gut and liver injury.
🗃️ See comments section for reference.
|
|
|
Scooped by
Gilbert C FAURE
March 27, 5:16 AM
|
Asthme : prévention, vaccination… et entretien pharmaceutique
On parle souvent des risques infectieux chez les patients asthmatiques — grippe, pneumocoque — et de l’importance de la vaccination pour éviter des exacerbations sévères.
Mais un levier reste encore sous-exploité : l’entretien asthme.
👉 Un moment clé pour évaluer le contrôle de la maladie
👉 Un espace pour vérifier l’observance et la bonne utilisation des dispositifs
👉 Une opportunité pour sensibiliser à la prévention, notamment vaccinale
👉 Un point de contact régulier dans le parcours de soins
C’est un dispositif puissant… à condition qu’il soit réellement activé.
Aujourd’hui, le défi n’est plus seulement d’informer, mais de structurer et de systématiser ces interactions.
C’est là que la technologie peut faire la différence.
Chez Apodis, nous travaillons à :
✔️ Identifier les patients éligibles à l’entretien asthme
✔️ Aider les pharmaciens à structurer leurs interventions
✔️ Intégrer la prévention vaccinale dans chaque échange
✔️ Suivre l’impact dans le temps
L’entretien asthme ne doit plus être une opportunité ponctuelle, mais un véritable outil de pilotage du parcours patient.
Et si chaque passage en pharmacie devenait un point d’activation de la prévention ?
|
|
Scooped by
Gilbert C FAURE
March 26, 4:12 AM
|
#Immunology | 𝗧 𝗖𝗲𝗹𝗹𝘀 𝗣𝗼𝘀𝗶𝘁𝗶𝗼𝗻 𝗧𝗵𝗲𝗺𝘀𝗲𝗹𝘃𝗲𝘀 𝗶𝗻 𝗟𝗼𝘄-𝗢𝘅𝘆𝗴𝗲𝗻 𝗭𝗼𝗻𝗲𝘀 𝘁𝗼 𝗖𝗼𝗺𝗺𝗮𝗻𝗱 𝗟𝘂𝗻𝗴 𝗗𝗲𝗳𝗲𝗻𝘀𝗲 | University of Basel researchers led by Jean de Lima found that specialized helper T cells migrate to oxygen-scarce edges of immune hubs during lung infection. There, they produce the so-called HIF-1α protein and release interleukin-21, directing macrophages, B cells, and natural killer cells into coordinated responses against respiratory pathogens. Using advanced imaging in influenza-infected mice and inducible knockout models, the team mapped how these cells position at hub boundaries to orchestrate defense networks.
The findings show tissue-resident immune hubs function as command centers for on-site protection rather than antibody factories. Professor Carolyn King's group validated the mechanism across secondary influenza infections and lung cancer models, showing broad therapeutic potential. This breakthrough enables design of inhalable vaccines that build immune defense directly in airways where viruses enter, potentially transforming respiratory disease prevention. The spatial coordination strategy also opens perspectives for tissue-targeted therapies that use the body's natural positioning systems to strengthen local immune responses at infection sites.
👇 Learn more & read the original publication: link in the comments 👇
🇨🇭 Follow #ScienceSwitzerland for the latest news and emerging trends on Swiss science, technology, education, and innovation >> swissinnovation.org
Follow us >> Science-Switzerland
#Science | #Education | #Research | #Innovation
|
|
Scooped by
Gilbert C FAURE
March 18, 6:49 AM
|
Pre-eclampsia is often described as a disease of the placenta, but at its core it is also a disease of immune balance. When the maternal immune system becomes overactivated, the delicate vascular architecture of the placenta can begin to fail.
Recent data showing rising pre-eclampsia rates after the pandemic has prompted renewed interest in immune-vascular triggers. If spike protein interacts with macrophages and endothelial cells in the placenta, understanding that mechanism could become a crucial piece of the puzzle in protecting maternal and fetal health.
========================================================
Your Gut Readiness Assessment
https://lnkd.in/ezPm4wht
========================================================
You can also find us Here:
Substack: https://lnkd.in/ejXM5s68
Videos: https://lnkd.in/g3vJ8uXx
Courses: https://lnkd.in/ecmtMKhz
Rumble: https://lnkd.in/gG_HmbdN
#covid #medicine #research
|
|
Scooped by
Gilbert C FAURE
March 9, 5:19 AM
|
Oral Vaccines Are Moving From Research to Reality
The global vaccine landscape may be entering a new phase — and oral vaccines are quickly becoming one of the most discussed innovations in infectious disease prevention.
In recent industry conversations across biotech and pharmaceutical research communities in the U.S., more attention is shifting toward oral vaccine platforms. Unlike traditional injections, oral vaccines could simplify distribution, improve patient compliance, and make large-scale immunization campaigns far more accessible — especially in regions where healthcare infrastructure is limited.
Several biotechnology companies are now accelerating research around oral delivery systems, mucosal immunity, and next-generation vaccine platforms. The idea is not only to prevent disease more effectively, but also to rethink how vaccines are manufactured, distributed, and administered globally.
For pharmaceutical companies and healthcare systems, this shift could represent more than just a scientific breakthrough. It may reshape public health logistics, vaccine accessibility, and global pandemic preparedness in the coming decade.
The question many people in the industry are asking now is:
If oral vaccines become widely scalable, could they fundamentally change the way the world approaches infectious disease prevention?
Curious to hear perspectives from people working across biotech, healthcare, and public health.
#Biotechnology #Vaccines #OralVaccines #PharmaceuticalIndustry #InfectiousDiseases
#DrugDevelopment #BiotechInnovation #GlobalHealth #HealthcareInnovation #ClinicalResearch
#LifeSciences #PublicHealth #BiotechInvesting #FutureOfMedicine #MedicalInnovation
|
|
Scooped by
Gilbert C FAURE
February 23, 4:27 AM
|
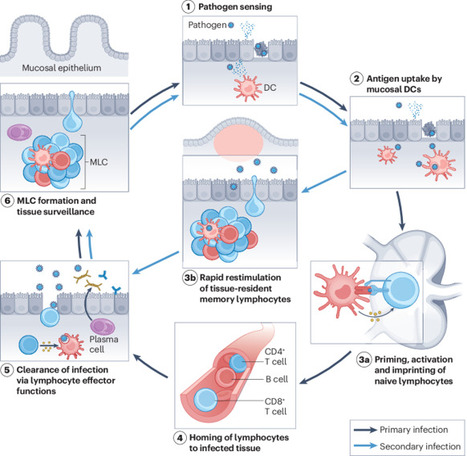
A very insightful review from the Akiko Iwasaki's group on the potential to harness mucosal immunity in next-generation vaccine development.
While the rapid deployment of intramuscular mRNA vaccines was a landmark achievement in preventing severe COVID-19, intramuscular shots often fall short of providing sterilizing immunity.
Mucosal immunity represents a particularly promising avenue for improving vaccines against respiratory viruses, as it enables immune protection to be established directly at the site of viral entry and early replication.
Mucosal tissues, such as the respiratory tract, host locally regulated specialized immune cells that are functionally and spatially distinct. Reduction of infection and transmission requires engaging the mucosal immune response: a coordinated process beginning with epithelial pathogen sensing and culminating in the establishment of tissue-resident memory T (TRM) and B (BRM) cells, alongside robust local secretory IgA (SIgA) production. Unlike systemic IgG, nasal SIgA has demonstrated superior virus-neutralizing activity and greater breadth against antigenically drifted variants.
A promising strategy for advancing vaccine design is the heterologous prime-boost approach. Research suggests that intramuscular priming (to establish peripheral memory pools) followed by an intranasal boost can effectively "pull" memory cells to the respiratory mucosa.
However, the so-called 'mucosal' vaccines requires navigating complex physiological constraints, such as the mucociliary clearance system and the anionic mucus layer.
Moreover, the regulatory path for mucosal vaccines is primarily hindered by the lack of validated correlates of protection, making it difficult to predict efficacy and guide clinical trial designs. Additionally, the anatomical proximity of the nasal mucosa to the central nervous system necessitates rigorous safety evaluations to prevent neuro-olfactory spillover or unintended neuro-inflammation.
https://lnkd.in/eb5ZUMY4
|
|
Scooped by
Gilbert C FAURE
February 20, 3:00 AM
|
Nasal spray vaccine could ‘replace multiple jabs every year’Bookmark popoverRemoved from bookmarksClose popoverScientists at Stanford Medicine have developed a universal vaccine formula, tested on mice, that offers broad protection against various respiratory threats. The vaccine, delivered as a nasal spray, could protect against cold, flu, Covid, allergies, respiratory viruses, sepsis-causing bacteria, and even house dust mites. It works by mimicking the signals immune cells use to communicate during an infection, rather than targeting specific parts of a pathogen. If developed for humans, this vaccine could replace multiple annual jabs for winter respiratory infections and potentially protect against new pandemic bugs. While lead author Dr Bali Pulendran estimates human availability within five to seven years, other experts caution that a truly universal vaccine is still some way off due to safety considerations and the diversity of the human population.
|
|
Scooped by
Gilbert C FAURE
February 16, 6:27 AM
|
Une avancée importante en recherche vaccinale pour les 6 mois à 5 ans
Une équipe dirigée par Guy Boivin, professeur au Département de pédiatrie et chercheur au Centre de recherche du CHU de Québec – Université Laval, a développé un vaccin expérimental administré par voie intranasale afin de protéger les jeunes enfants contre deux virus respiratoires majeurs : le métapneumovirus humain et le virus respiratoire syncytial (VRS). Ces deux agents infectieux sont responsables chaque année de nombreuses bronchiolites et pneumonies chez les jeunes enfants.
Les premières études menées sur des modèles animaux montrent des résultats très encourageants.
Cette avancée repose sur une plateforme vaccinale qui permet d’intégrer rapidement des éléments de différents virus pour créer de nouveaux candidats vaccins.
Découvrez tous les détails :
|
|
Scooped by
Gilbert C FAURE
February 10, 4:48 AM
|
|
|
Scooped by
Gilbert C FAURE
February 4, 6:39 AM
|
Ever had a client ask: "Why does my kitten need to come TWO times for vaccines? Can't we just do it all at once?"
Let me break down the science in a way that might change how it act
🧬 The "Goldilocks Problem" of Maternal Immunity
Kittens are born with almost NO immunity from their mother during pregnancy. Unlike humans, cats have a special type of placenta that blocks antibody transfer before birth. Instead, 90-95% of protective antibodies come through colostrum in those critical first 16 hours of life (Claus et al., 2006).¹
But here's where it gets tricky...
These maternal antibodies are both a blessing and a curse:
✅ They protect vulnerable kittens from deadly diseases
❌ But they ALSO attack vaccine antigens, preventing the kitten from building their own immunity
This creates what scientists call the "window of susceptibility", a period where kittens are:
-Too vulnerable to fight off real infections
-Yet unable to respond to vaccines
Consider these exposure risks for "indoor-only" cats:
-Panleukopenia virus survives for YEARS in the environment and can be tracked indoors on shoes and clothing
-Multi-cat households where ONE cat goes outside creates risk for ALL cats
Here's what evidence-based feline vaccination looks like in #2026:
For Kittens: → Start at 6-8 weeks, continue every 2-4 weeks until 16-20 weeks → Core vaccines: FPV, FHV-1, FCV → FeLV for ALL kittens (remember that age-resistance curve!) → Rabies at 12-16 weeks → yearly booster
For Adult High-Risk Cats: → Annual booster of Core and Rabies
What challenges do you face while Vaccination?
#mianpetsandvets #VeterinaryMedicine #FelineHealth #VetMed #CatVaccination #VeterinaryEducation #CatsOfLinkedIn
|
|
Scooped by
Gilbert C FAURE
January 24, 7:33 AM
|
Sign in or join now to see posts like this one and more.
|
|
Scooped by
Gilbert C FAURE
January 12, 9:07 AM
|
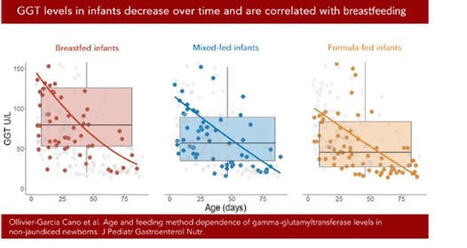
Une belle façon de démarrer 2026 ! Notre étude sur les taux de gamma-GT sanguins chez les nourrissons allaités vient d’être publiée dans le Journal of Pediatric Gastroenterology and Nutrition (JPGN). Ce travail, issu de la thèse d’Audrey Ollivier-Garcia Cano, a été mené en collaboration avec Marion Marlinge, Paul Guerry et Aurélie MORAND .
En médecine vétérinaire, le taux de gamma-GT est utilisé comme marqueur du transfert d’immunoglobulines via le colostrum chez les bovins (les IgG ne passant pas la barrière placentaire). Certaines études suggéraient par ailleurs que le lait maternel est riche en gamma-GT. Nous avons donc mené une étude rétrospective chez des nourrissons ayant eu un bilan hépatique et consultant aux urgences pédiatriques, en excluant ceux présentant une infection ou un ictère.
Les gamma-GT sériques étaient significativement plus élevées chez les enfants allaités (101 UI/l) que chez les non-allaités (64 UI/l), avec un niveau intermédiaire (77 UI/l) pour l’allaitement mixte. Nous avons également confirmé la diminution des gamma-GT avec l’âge.
Ces résultats soulignent l’importance d’interpréter les dosages de gamma-GT en fonction du mode d’allaitement. Ils rappellent aussi la richesse des approches pluridisciplinaires.
https://lnkd.in/dEnBgETK
|
|
Scooped by
Gilbert C FAURE
January 8, 1:43 PM
|
The gut virome is a complex ecosystem characterized by the interplay of diverse viral entities, predominantly bacteriophages and eukaryotic viruses. The gut virome has a critical role in human health by shaping microbial community profiles, modulating host immunity and influencing metabolic processes. Different viral metagenomics approaches have revealed the remarkable diversity of the gut virome, showing individual-specific patterns that evolve over time and adapt dynamically to environmental factors. Perturbations in this community are increasingly associated with chronic immune and inflammatory conditions, metabolic disorders and neurological conditions, highlighting its potential as a diagnostic biomarker and therapeutic target. The early-life gut virome is particularly influential in establishing lifelong health trajectories through its interactions with diet, immune pathways and others, thereby contributing to inflammatory and metabolic regulation. This Review synthesizes current knowledge of gut virome composition, dynamics and functional relevance, critically evaluating evidence distinguishing causal from correlative roles in disease pathogenesis. The interactions of the virome with other microbiome components and host immunity are examined, and emerging translational applications, including phage therapy and biomarker development, are discussed. Integrating these insights while acknowledging methodological challenges provides a comprehensive framework for understanding the complex roles of the gut virome in health and disease. The gut virome is a complex ecosystem and has a critical role in human health. This Review outlines gut virome composition and functional relevance, and its role in human health and disease. Methodological challenges in advancing our knowledge of the gut virome are also discussed.
|
|
Scooped by
Gilbert C FAURE
January 5, 4:21 AM
|
IgA friends at mucossl level and foes at systemic level… a new proof…
|