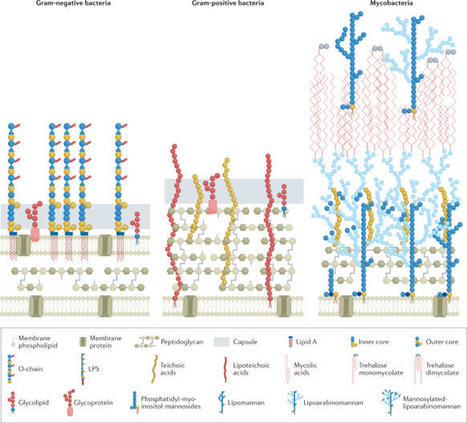
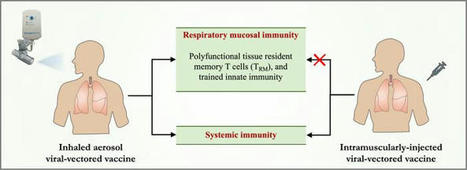
Your new post is loading...
Your new post is loading...

|
Scooped by
Gilbert C FAURE
December 17, 2022 2:41 AM
|
The new knowledge will allow fish immunologists better understand the efficacy of aquaculture vaccines.
|
|
Scooped by
Gilbert C FAURE
November 9, 2022 1:29 PM
|
Polymeric IgA and IgM are transported across the epithelial barrier from plasma cells in the lamina propria to exert a function in the gut lumen as secretory antibodies. Many secretory antibodies are reactive with the gut bacteria, and mounting evidence suggests that these antibodies are important for the host to control gut bacterial communities. However, we have incomplete knowledge of how bacteria-reactive secretory antibodies are formed. Antibodies from gut plasma cells often show bacterial cross-species reactivity, putting the degree of specificity behind anti-bacterial antibody responses into question. Such cross-species reactive antibodies frequently recognize non-genome-encoded membrane glycan structures. On the other hand, the T cell epitopes are peptides encoded in the bacterial genomes, thereby allowing a higher degree of predictable specificity on the T cell side of anti-bacterial immune responses. In this Perspective, we argue that the production of bacteria-reactive secretory antibodies is mainly controlled by the antigen specificity of T cells, which provide help to B cells. To be able to harness this system (for instance, for manipulation with vaccines), we need to obtain insight into the bacterial epitopes recognized by T cells in addition to characterizing the reactivity of the antibodies. Secretory antibodies regulate intestinal bacteria, but their specificity and dependence on T cells have been matters of debate. This Perspective argues that bacteria-reactive antibodies are highly specific and that their production is under the control of T cells.
|
|
Scooped by
Gilbert C FAURE
October 12, 2022 10:22 AM
|
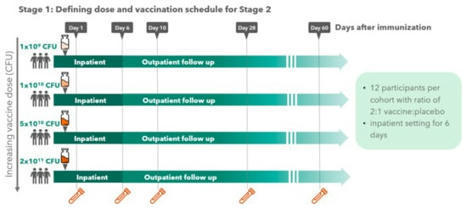
During the period of March 2019 to February 2021, we enrolled 36 BCG-vaccinated healthy adults between 18 and 55 years of age at McMaster University Medical Centre. Four participants were excluded (2 withdrew consent and 2 were withdrawn before vaccination because they were unable to comply with the study visit requirements) and 1 did not complete any follow-up visits after vaccination because of COVID restrictions. Thirty-one participants completed the study: 11 in the low-dose (LD) aerosol group, 11 in the high-dose (HD) aerosol group, and 9 in the i.m. group (Figure 1). The demographic and baseline characteristics of the study participants were similar among study groups (Table 1). Figure 1Trial profile. PPD, purified protein derivative; LD, low-dose aerosol; HD, high-dose aerosol; i.m.-intramuscular injection; PFU, plaque-forming unit. Table 1Demographics of participants. Characterization of inhaled aerosol delivery method and aerosol droplets using Aeroneb Solo device. The Aeroneb Solo Micropump was selected to be part of the device set up for aerosol generation and delivery in our study (Supplemental Figure 1; supplemental material available online with this article; https://doi.org/10.1172/jci.insight.155655DS1). A fill volume (FV) of 0.5 mL in the nebulizer was determined to be optimal for vaccine delivery in saline. Subjects completed inhalation of this volume containing the vaccine via tidal breathing in approximately 2.5 minutes (Table 2). The emitted dose (ED) of vaccine available at the mouth was found to be approximately 50% of the loaded dose in the nebulizer (Table 2). The majority of aerosol droplets containing the vaccine were < 5.39 μm (85%), or between 2.08 and 5.39 μm in diameter, conducive to vaccine deposition in major airways. Thus, the amount of aerosol available at the mouth and subsequently deposited in the lung was 42.5% (16). The estimated rate of viable vaccine from aerosol droplets generated by the nebulizer was 17.4%. The dose loaded in the nebulizer for aerosol inhalation was, thus, corrected according to the estimated losses of vaccine within the device. Table 2Characterization of aerosol device and aerosol droplets. Safety of inhaled aerosol and i.m.-injected AdHu5Ag85A vaccine. Both LD (1 × 106 PFU) and HD (2 × 106 PFU) of AdHu5Ag85A administered by aerosol inhalation or the i.m. injection were safe and well tolerated. Respiratory adverse events were infrequent, mild, transient, and similar among groups (Table 3). I.m. injection was associated with a mild local injection site reaction in 2 participants. Systemic adverse events were also infrequent, mild, transient, and similar among groups (Table 3). One participant who received LD aerosol vaccine developed genital lesions consistent with primary HSV-1 infection the day following the week-2 bronchoscopy, and this condition resolved without complication with oral valacyclovir; one participant developed plantar fasciitis on day 13 following vaccination, which was attributed to mechanical strain and resolved with acetominophen. There were no grade 3 or 4 adverse events reported, nor any serious adverse events. Table 3Adverse events. There were no clinically significant abnormalities of laboratory tests at weeks 2, 4, and 12 following vaccination. Follow-up respiratory functional determinations forced expiratory volume in the first second (FEV1)and forced vital capacity (FVC) were similar to baseline values in all participants across all 3 groups (Figure 2, A and B). Figure 2Respiratory function and bronchoalveolar cellular responses following aerosol or intramuscular vaccination. (A and B) Lung function was assessed as FEV1 and FVC at baseline and 2 weeks after LD aerosol (n = 11), HD aerosol (n = 11), or i.m. (n = 9) vaccination. (C–F) Frequencies of differential cells including macrophages, lymphocytes, neutrophils, and epithelial cells in BALF from LD aerosol, HD aerosol, and i.m. vaccine cohorts. Violin plots show the median and quartiles. Data in dot plots are expressed as the mean value (horizontal line) with 95% CI. Wilcoxon matched pairs signed-rank test was used to compare various time points with baseline values within the same vaccination group. Bronchoscopy and bronchoalveolar lavage were generally well tolerated in all participants. As expected, in some participants, the procedures were associated with mild cough, sore throat, low-grade fever, headache, and a transient drop in FEV1. The appearance of the bronchial mucosa was judged as normal in all participants at each time point. Adequate bronchoalveolar lavage fluid (BALF) volumes were obtained following bronchoalveolar lavage, and on average, 10 to 20 million total cells were obtained. Aerosol AdHu5Ag85A vaccination induces robust and sustainable Th1 responses in the airway. Bronchoalveolar lavage was obtained successfully at baseline and at 2 and 8 weeks after vaccination from all participants with a median return volume of 87.5 mL (IQR, 72.5–98) from a total of 160 mL saline instilled and a median total cell number of 0.14 million/mL BALF (IQR, 0.1–0.2). Cellularity in the airway significantly increased 2 weeks after both LD and HD aerosol vaccination, and in the LD aerosol group, cellularity remained significantly heightened up to 8 weeks after vaccination compared with baseline (Figure 2C). Both LD and HD aerosol vaccination led to a transient reduction in airway macrophages, but the lymphocyte counts significantly increased only in LD cohort (Figure 2, D and E). Importantly, both neutrophils and epithelial cells in the airway remained either absent or unaltered following aerosol vaccination (Figure 2, D and E), indicating no significant airway inflammation except vaccine-induced lymphocytic responses. In comparison, there were no marked changes in total cellularity and any leukocyte subsets in the airway after i.m. vaccination (Figure 2, C and F). Evaluation of Th1 responses in the airways (BALF) cells was performed by intracellular cytokine immunostaining and flow cytometry (the gating strategy shown in Supplemental Figure 2). It showed that both LD and HD aerosol AdHu5Ag85A markedly increased Ag85A peptide pool–specific (Ag85A p. pool–specific) or reactive, IFN-γ–, TNF-α–, and/or IL-2–producing CD4+ T cells in the airways at 2 weeks after vaccination compared with the respective baseline responses (Figure 3, A–C). On average, the total Ag-specific cytokine-producing CD4+ T cells represented approximately 25% of all CD4+ T cells at 2 weeks after LD or HD aerosol, and they remained significantly elevated up to 8 weeks in the airways of LD cohort (Figure 3, A and B). In contrast, i.m. AdHu5Ag85A vaccination failed to induce Ag-specific CD4+ T cells in the airways (Figure 3A). Compared with CD4+ T cells, although the levels of airway CD8+ T cell responses were much smaller, they were significantly increased at both 2 and 8 weeks, particularly following LD aerosol vaccination (Figure 3D). Of interest, there was also a small but increased number of CD8+ T cells at 2 weeks after i.m. vaccination. Figure 3Induction of multifunctional T cells in the airways following aerosol or i.m. vaccination. (A) Frequencies of airway antigen–specific combined total-cytokine–producing CD4+ T cells at various time points in LD aerosol, HD aerosol, and i.m. cohorts. (B) Frequencies of airway single-cytokine–producing CD4+ T cells at various time points in LD aerosol cohort. (C) Frequencies of airway single-cytokine–producing CD4+ T cells at various time points in HD aerosol cohort. (D) Frequencies of airway antigen–specific combined total-cytokine–producing CD8+ T cells at various time points in LD aerosol, HD aerosol, and i.m. cohorts. (E) Representative dot plots of airways CD4+ and CD8+ T cells expressing IFN-γ, TNF-α, and IL-2 at wk2 from LD aerosol participants. (F) Frequencies of airways polyfunctional (triple/3+, double/2+, and single/1+ cytokine+) antigen-specific CD4+ and CD8+ T cells at various time points in LD aerosol group. (G) Median proportions displayed in pie chart of antigen-specific airways CD4+ and CD8+ T cells expressing a specific single or combination of 2 or 3 cytokines at various time points in LD aerosol group. Data in dot plots are expressed as the mean value (horizontal line) with 95% CI. Box plots show mean value (horizontal line) with 95% CI (whiskers), and boxes extend from the 25th to 75th percentiles. Wilcoxon matched pairs signed-rank test was used to compare various time points with baseline values within the same vaccination group. Analysis of polyfunctionality of vaccine-activated CD4+ and CD8+ T cells reactive to Ag85A in the airways revealed that LD aerosol vaccination led to induction of a higher magnitude of CD4+ T cells that coexpressed IFN-γ, TNF-α, and IL-2 (3+) and any of 2 cytokines (2+) compared with those producing single cytokine (1+) (Figure 3, E and F). Importantly, polyfunctional CD4+ T cells remained significantly increased over the baseline up to 8 weeks. Similarly, LD aerosol vaccination also significantly increased the polyfunctional CD8+ T cells, particularly at 8 weeks, in the airways, though at a much lower overall magnitude compared with CD4+ T cells (Figure 3F). In comparison, HD aerosol vaccination led to significantly increased polyfunctional CD4+ but not CD8+ T cells reactive to Ag85A (Supplemental Figure 3, A and B). Given that the LD aerosol AdHu5Ag85A was consistently highly immunogenic, we profiled the polyfunctional CD4+ and CD8+ T cells in the airways of the LD cohort in greater detail. While at 2 weeks, a greater proportion of Ag85A-reactive CD4+ T cells were polyfunctional (IFN-γ+TNF-α+IL-2+, IFN-γ+TNF-α+, or IFN-γ+IL-2+), with some of them also being single cytokine producers, at 8 weeks, the vast majority of them (>95%) became polyfunctional (Figure 3G). In comparison, most of the CD8+ T cells at 2 weeks were single-cytokine producers (TNF-α+), but at 8 weeks, the majority of them turned to be polyfunctional, mostly being IFN-γ+TNF-α+ (Figure 3G). Since the trial participants were previously BCG vaccinated, we examined the overall T cell reactivity to stimulation with multimycobacterial antigens. We found considerable CD4+ T cells present in the airways to be reactive to a cocktail of mycobacterial antigens even prior to vaccination, and they remained unaltered after LD, HD, or i.m. vaccination (Supplemental Figure 3C). These BCG-specific CD4+ T cells in the airways of LD group were mostly polyfunctional (Supplemental Figure 3, D and E). Similarly, small numbers of preexisting BCG-specific CD8+ T cells in the airways were not altered by aerosol or i.m. vaccination (Supplemental Figure 3F). The above data suggest that inhaled aerosol, but not i.m., AdHu5Ag85A vaccination can induce robust antigen-specific T cell responses within the respiratory tract. Furthermore, a LD (1 × 106 PFU) aerosol vaccination is superior to a HD (2 × 106 PFU) aerosol in inducing robust and sustainable respiratory mucosal immunity. The mucosal responses induced by AdHu5Ag85A vaccine are predominantly polyfunctional CD4+ T cells in nature, with some levels of polyfunctional CD8+ T cells. The preexisting CD4+ T cells of multimycobacterial antigen specificities in the airway of BCG-vaccinated trial participants were not significantly impacted by AdHu5Ag85A aerosol vaccination. Aerosol vaccination induces airway TRM expressing the lung-homing molecule α4β1 integrin. Lung tissue TRM are critical to protective mucosal immunity (17). Hence, we next determined whether antigen-specific T cells induced by aerosol AdHu5Ag85A vaccination were of tissue-resident memory phenotype and compared them with those induced by i.m. vaccination. BALF cells obtained before and at select time points after vaccination were stimulated with Ag85A p. pool and immunostained for coexpression of 2 key TRM surface markers CD69 and CD103 by antigen-specific IFN-γ–producing CD4+ or CD8+ T cells (Figure 4A). Marked increases in Ag85A-specific IFN-γ+ CD4+ and CD8+ T cells coexpressing CD69 and CD103 were seen only in the airway of LD and HD aerosol vaccine groups and not in i.m. group (Figure 4, B and C). Although TRM increases at 8 weeks after aerosol vaccination, compared with the baseline, were only marginally statistically significant (95% CI) probably due to small sample sizes, remarkable proportions of Ag85A-specific CD4 T cells (~20%) and CD8+ T cells (~54%) present in the airways of aerosol vaccine groups were TRM (Figure 4D). As expected, there was no detectable antigen-specific TRM in the peripheral blood before and after vaccination. Figure 4Induction of airway tissue TRM following aerosol or i.m. vaccination. (A) Representative dot plots of airway antigen–specific CD4+ and CD8+ TRM at wk2 in LD aerosol participants. (B) Frequencies of airway antigen–specific IFN-γ+CD4+ TRM coexpressing CD69 and CD103 surface markers at various time points in LD aerosol, HD aerosol, and i.m. vaccine cohorts. (C) Frequencies of airway antigen–specific IFN-γ+CD8+ TRM coexpressing CD69 and CD103 at various time points in LD aerosol, HD aerosol, and i.m. vaccine cohorts. (D) Comparison of frequencies of airway antigen–specific CD4+ and CD8+ TRM coexpressing CD69 and CD103 at 8 weeks after LD and HD aerosol vaccination. (E) Representative dot plots of peripheral blood antigen–specific IFN-γ+CD4+ T cells expressing CD49d at wk2 from LD aerosol participants, and frequencies of circulating antigen-specific CD4+ T cells expressing CD49d at various time points in LD aerosol, HD aerosol, and i.m. vaccine cohorts. (F) Representative dot plots of airway antigen–specific IFN-γ+CD4+ T cells expressing CD49d at wk2 from LD aerosol participants, and frequencies of airway antigen–specific CD4+ T cells expressing CD49d at various time points in LD aerosol, HD aerosol, and i.m. vaccine cohorts. (G) Comparison of frequencies of airway antigen–specific IFN-γ+CD4+ T cells coexpressing CD49d at the peak time point in LD aerosol, HD aerosol, and i.m. vaccine cohorts. Data in dot plots are expressed as the mean value (horizontal line) with 95% CI. Wilcoxon matched pairs signed-rank test (B, C, E, and F) was used to compare various time points with baseline values within the same vaccination group. Mann-Whitney U test (D and G) was used when comparing between vaccination groups. We also studied T cell surface expression of α4β1 integrin (VLA-4; or CD49d for α4), known to be expressed on memory CD4+ T cells in human airways (18). Since CD49d may be involved in the homing of circulating T cells to the airway, we first examined CD49d expression on Ag85A-specific CD4+ T cells in the circulation. There were small but significantly increased frequencies of circulating CD49d-expressing IFN-γ+CD4+ T cells, particularly at 2 weeks following LD or HD aerosol vaccination (Figure 4E). In comparison, there were much greater frequencies of CD49d-expressing IFN-γ+CD4+ T cells (out of total CD4+ T cells) in the airways induced by aerosol vaccination (Figure 4F), compared with their frequencies in the circulation (Figure 4E) and in contrast with the lack of such T cells in the airways of i.m. group (Figure 4G). In fact, the majority of Ag85A-specific CD4+ T cells in the airways of LD and HD groups expressed CD49d (57% and 74%, respectively). The data indicate that aerosol AdHu5Ag85A vaccination, but not i.m. route of vaccination, is uniquely capable of inducing antigen-specific T cells in the airways endowed with respiratory mucosal homing and TRM properties. Since, besides mucosal adaptive immunity, respiratory delivery of AdHu5Ag85A vaccine in experimental animals induced a trained phenotype in airway macrophages (6, 19), we examined whether aerosol vaccination could also alter the immune property of human alveolar macrophages (AM). To this end, we elected to examine the transcriptomics of BALF cells obtained from 5 participants before (week 0 [wk0]) and after (week 8 [wk8]) LD aerosol vaccination. Before RNA isolation, the cells, upon revival from frozen stock, were enriched for AM and cultured with or without stimulation with M. tuberculosis lysates and transcriptionally profiled by RNA-Seq analysis. Principal component analysis (PCA) revealed that unstimulated and stimulated AM populations were separated away from each other (Figure 5A). We then identified the differentially expressed genes (DEGs) by comparing wk0-stimulated (Group 3–stimulated) and wk8-stimulated (Group 4–stimulated) AM with respective unstimulated AM (wk0/Group 1) and wk8/Group 2). A total of 2726 genes was differentially expressed upon stimulation in pairwise analysis, of which 1667 genes (61%) were shared between the baseline (wk0) Group 3/Group 1 and aerosol vaccine (wk8) Group 4/Group 2 (Figure 5B). As expected, the shared genes were significantly enriched in biological processes associated with immune response and regulation of cell death (Figure 5C). Furthermore, by pairwise analysis, we identified 191 and 426 genes uniquely upregulated and downregulated, respectively, in stimulated aerosol (Group 4) AM (Figure 5D). The uniquely upregulated genes in stimulated wk8 aerosol AM showed enrichment in a number of biological processes including response to anoxia (OXTR, CTGF), inflammatory response to antigenic stimuli (IL-2RA, IL-1B, IL-20RB), tyrosine phosphorylation of STAT protein (IFN-γ, F2R, OSM), regulation of IL-10 production (CD83, IRF4, IL-20RB, IDO1), response to IL-1 (RIPK2, SRC, IRAK2, IL-1R1, XYLT1, RELA), and histone demethylation (KDM6B, KDM5B, KDM1A, KDM7A, JMJD6; Figure 5E). In comparison, the uniquely downregulated genes in wk8 aerosol AM did not appear significantly enriched for any biological processes. These data suggest that LD aerosol vaccination leads to persisting transcriptional changes in airway-resident AM poised for defense responses. Figure 5Transcriptomic analysis of alveolar macrophages (AM) following LD aerosol vaccination. (A) Principal component analysis (PCA) of gene expression in AM obtained before (wk0) and after (wk8) LD aerosol vaccination cultured with (S) or without (US) stimulation. (B) Venn diagram comparing all DEGs in pairwise comparison. (C) Significantly enriched functional categories of biological processes by GO associated with DEGs shared between the baseline (wk0) Group 3/1 and aerosol vaccine (wk8) Group 4/2. (D) Venn diagram comparing up- and downregulated DEGs in pairwise comparison. Heatmap shows DEG uniquely up- and downregulated, in stimulated aerosol (Group 4) AM. (E) Significantly enriched functional categories of biological processes by GO associated with uniquely upregulated DEGs in stimulated aerosol (Group 4) AM. Statistical differences in functional categories of biological processes was performed using BINGO plugin, which uses a hypergeometric test with Benjamini-Hochberg FDR correction. Both aerosol and i.m. vaccination induce systemic Th1 responses. Assessment of overall antigen-specific reactivity of T cells in the circulation before and after vaccination by using whole blood samples incubated with Ag85A peptides indicated that both aerosol, particularly LD aerosol, and i.m. AdHu5Ag85A vaccination induced significant systemic immune responses, as shown by raised IFN-γ, TNF-α, and IL-2 levels in plasma (Figure 6, A–C). AUC analysis, which reflects the overall magnitude of responses, did not differ between LD aerosol and i.m. groups in cytokine production in response to Ag85A p. pool stimulation (IFN-γ, P = 0.0910; TNF-α, P = 0.6207; IL-2, P = 0.8703). However, i.m. vaccine–induced systemic T cell responses appeared to remain significantly increased over a longer duration (Figure 6C). In comparison, the HD aerosol group had significantly lower IFN-γ production than i.m. group (AUC compared with i.m., P = 0.0117) whereas they did not differ from each other in the production of TNF-α and IL-2 (Figure 6, B and C). Figure 6Induction of antigen-specific T cell responses in the peripheral blood following aerosol or intramuscular vaccination. (A–C) Antigen-specific cytokine production in whole blood culture at various time points after LD aerosol, HD aerosol, and i.m. vaccine groups. The measurements were subtracted from unstimulated control values. (D) Frequencies of peripheral blood antigen–specific combined total-cytokine-producing CD4+ T cells at various time points in LD aerosol, HD aerosol, and i.m. cohorts. (E) Frequencies of peripheral blood polyfunctional (triple/3+, double/2+, and single/1+ cytokine+) antigen-specific CD4+ T cells at various time points in LD aerosol, HD aerosol, and i.m. vaccine groups. (F) Frequencies of peripheral blood antigen–specific combined total-cytokine–producing CD8+ T cells at various time points in LD aerosol, HD aerosol, and i.m. groups. (G) Frequencies of peripheral blood polyfunctional (triple/3+, double/2+ and single/1+ cytokine+) antigen-specific CD8+ T cells at various time points in LD aerosol, HD aerosol, and i.m. groups. Data in dot plots are expressed as the mean value (horizontal line) with 95% CI. Box plots show mean value (horizontal line) with 95% CI (whiskers), and boxes extend from the 25th to 75th percentiles. Line graphs show median with IQR. Wilcoxon matched pairs signed-rank test (A, B, E, and G) was used to compare various time points with baseline values within the same vaccination group. Mann-Whitney U test (D and F) was used when comparing vaccination groups. Further examination of relative activation of CD4+ and CD8+ T cells by aerosol and i.m. vaccinations using intracellular cytokine staining (ICS) revealed that, compared with the respective baseline, aerosol vaccination activated the circulating Ag85A-specific CD4+ T cells to significant levels, while i.m. vaccination moderately increased such responses (Figure 6D). However, the overall magnitude of responses did not differ significantly between aerosol and i.m. groups (AUC: LD aerosol, P = 0.1961; HD aerosol, P = 0.3545 compared with i.m.). Both LD/HD aerosol and i.m. vaccination also significantly increased Ag85A-specific polyfunctional CD4+ T cells coexpressing 3 (3+) or any 2 (2+) cytokines in the circulation (Figure 6E). Consistent with the airway Ag85A-specific CD4+ T cell responses (Figure 3, A–C) in both LD and HD aerosol groups, circulating polyfunctional CD4+ T cells also generally peaked at 2 weeks after vaccination and remained significantly increased up to 8 weeks (Figure 6E). In comparison, 3+ polyfunctional CD4+ T cells in the i.m. group significantly increased at 4 weeks and remained increased up to 8 weeks (Figure 6E). The overall magnitude of circulating 3+ polyfunctional CD4+ T cells in the i.m. group was, however, significantly higher than those in aerosol groups (AUC: LD aerosol, P = 0.0144; HD aerosol, P = 0.0393 compared with i.m.). Circulating 2+ polyfunctional CD4+ T cells did not differ significantly between these groups (AUC not significantly different). Consistent with our previous observation (8), besides its activating effects on circulating CD4+ T cells, i.m. vaccination also significantly increased Ag85A-specific CD8+ T cells up to 16 weeks (Figure 6F). By comparison, aerosol vaccination minimally induced such CD8+ T cells in the circulation (Figure 6F). Compared with circulating CD4+ T cells (Figure 6E), similar to the overall kinetics of total-cytokine+ CD8+ T cells (Figure 6F), circulating Ag85A-specific polyfunctional CD8+ T cells peaked behind the peak CD4+ T cell responses in all vaccine groups (Figure 6G). The kinetics of polyfunctional profiles of circulating CD4+ T cells were further examined in greater detail with a focus on the LD aerosol vaccine group and its comparison with the i.m. group. There existed considerable differences in the polyfunctional profile of circulating Ag85A-specific CD4+ T cells between LD aerosol and i.m. groups (Supplemental Figure 4A). In the LD aerosol group, the proportion of IFN-γ+TNF-α+IL-2+ progressively shrank, and at 16 weeks, approximately 75% of the population were TNF-α+IL-2+ and IFN-γ+TNF-α+ together with single TNF-α+ CD4+ T cells. In comparison, in the i.m. group, the proportion of IFN-γ+TNF-α+IL-2+ progressively expanded, constituting approximately 75% of the population at 16 weeks (Supplemental Figure 4A). Upon examination of circulating BCG-specific CD4+ T cells (reactive to M. tuberculosisCF+ rAg85A stimulation), we found that they were not strikingly increased in aerosol and i.m. vaccine groups, although the trend was higher in i.m. group (Supplemental Figure 4, B and C), and AUC values did not differ significantly between aerosol and i.m. groups (LD aerosol, P = 0.0870; HD aerosol, P = 0.2666, compared with i.m.). However, LD and HD aerosol vaccination had a significant enhancing effect on the polyfunctionality of preexisting circulating BCG-specific CD4+ T cells (Supplemental Figure 4C). Similar to BCG-specific circulating CD4+ T cells (Supplemental Figure 4B), BCG-specific circulating CD8+ T cells were not significantly increased by either aerosol or i.m. vaccination (Supplemental Figure 4D). These data indicate that, besides markedly induced mucosal T cell immunity (Figures 3 and 4), respiratory mucosal vaccination via inhaled aerosol, particularly LD aerosol, can also induce systemic polyfunctional CD4+ T cell responses, similar to i.m. route of vaccination in previously BCG-vaccinated humans. Preexisting and vaccine-induced anti-AdHu5 Ab in the circulation and airways. The high prevalence of circulating preexisting antibodies (Ab) against AdHu5 in human populations may negatively impact the potency of AdHu5-vectored vaccines following i.m. administration (20). However, little is known about its effect on the potency of AdHu5-vectored vaccine delivered via the respiratory mucosa. To address this question, we first examined the levels of AdHu5-specific total IgG in the circulation and airways (BALF) before and after vaccination (wk0 versus wk4 in circulation; wk0 versus wk8 in BALF). In keeping with our previous findings (8), there were significant levels of preexisting circulating AdHu5-specific total IgG in most of the trial participants (1 × 104 to 1 × 105), and the levels were comparable between the groups (using Kruskal-Wallis test P = 0.2048; Table 4). These titres significantly increased after HD aerosol or i.m. AdHu5Ag85A vaccination but not after LD aerosol vaccination. In comparison, preexisting levels of anti-AdHu5 total IgG in the airways were 1 to 1.5 log less than the levels in the circulation and were comparable between groups (using Kruskal-Wallis test, P = 0.2048). Of interest, LD and HD aerosol, as well as i.m. vaccination, did not alter the preexisting anti-AdHu5 total IgG levels in the airways (Table 4), but the data from HD aerosol and i.m. groups should be interpreted with caution due to the small sample size at 8 weeks. Table 4Anti-Ad5 antibody titers before and after LD aerosol, HD aerosol, and i.m. vaccination. Because the total anti-AdHu5 Ab titres may not always correlate with AdHu5-neutralizing capacity in the circulation (8), we further assessed the AdHu5-neutralizing Ab (nAb) titres before and after vaccination in the circulation and airways by using a bioassay. The preexisting AdHu5 nAb titres in the circulation were comparable between groups (using Kruskal-Wallis test, P = 0.3588) with 27%, 54%, and 66% of participants in LD, HD, and i.m. groups having > 1 × 102 AdHu5 nAb titres, respectively. Of interest, while i.m. vaccination with AdHu5Ag85A significantly increased the circulating AdHu5 nAb titers by an average of 1.5 logs, LD or HD aerosol vaccination had no such effect (Table 4). On the other hand, similar to total anti-AdHu5 IgG levels, preexisting AdHu5 nAb titers in the airways were ~1 log less than those in the circulation (Table 4). Of importance, 63%, 36%, and 33% of participants in LD, HD., and i.m. groups, respectively, had no detectable baseline AdHu5 nAb titers in their airways, which remained unaltered following vaccination (Table 4). We further found a significant positive correlation between AdHu5 nAb and total AdHu5 IgG titres both in the circulation and airways (Supplemental Figure 5, A and B). Given that many of the trial participants had moderate to significant levels of AdHu5 nAb titers in the circulation and ~50% of them also had a small but detectable level of preexisting AdHu5 nAb titres in the airways, we next examined whether such nAbs present in the airways and blood may have negatively impacted the immunopotency of LD aerosol and i.m. vaccination, respectively. To this end, the percentage of airways or blood with total-cytokine+ Ag85A-specific CD4+ T cells at the peak response time (2 weeks post-vaccination) for individual participants was plotted against corresponding preexisting AdHu5 nAb titres, and Spearman rank correlation test was performed. There was no significant correlation between preexisting airways AdHu5 nAb titers and the magnitude of vaccine-induced CD4+ (Supplemental Figure 5C) and CD8+ (Supplemental Figure 5D) T cell responses in the airways following LD aerosol vaccination. Of note, one participant who hardly responded to aerosol vaccine did have the highest neutralization titers in the cohort (Supplemental Figure 5, C and D). On the other hand, consistent with our previous observation (8), there was no significant correlation between preexisting circulating AdHu5 nAb titers and the magnitude of antigen-specific CD4+ (Supplemental Figure 5E) and CD8+ (Supplemental Figure 5F) T cell responses in the blood following i.m. vaccination. The above data suggest that, while there is high prevalence of preexisting circulating anti-AdHu5 nAb in humans enrolled in our study, most trial participants have either undetectable or very low levels of preexisting anti-AdHu5 nAb titers in the airways. I.m. AdHu5Ag85A vaccination increases AdHu5 nAb titers in the circulation, whereas aerosol vaccination does not do so either in the airways or in the circulation. Although the presence of AdHu5 nAb in the airways does not seem to have a significant impact on aerosol vaccine immunogenicity, the data should be interpreted with caution due to the small sample size and very few BALF samples with significant AdHu5 nAb titers.
|
|
Scooped by
Gilbert C FAURE
October 8, 2022 10:45 AM
|
Mucosal Vaccines for SARS-CoV-2: Scientific Gaps and Opportunities. Online registration by Cvent
|
|
Scooped by
Gilbert C FAURE
August 21, 2022 2:46 AM
|
Global immunization programmes have achieved some remarkable successes. In 1977, Frank Fenner's Commission declared smallpox to have been erad...
|
|
Scooped by
Gilbert C FAURE
August 18, 2022 5:11 AM
|
Analysis: a bivalent vaccine has been approved and research is being carried out into possible pan-coronavirus vaccines...
|
|
Scooped by
Gilbert C FAURE
August 13, 2022 11:08 AM
|
The mRNA vaccine platform has offered the greatest potential in fighting the COVID-19 pandemic owing to rapid development, effectiveness, and scalability to meet the global demand. There are many other mRNA vaccines currently being developed against different emerging viral diseases. As with the current COVID-19 vaccines, these mRNA-based vaccine candidates are being developed for parenteral administration via injections. However, most of the emerging viruses colonize the mucosal surfaces prior to systemic infection making it very crucial to target mucosal immunity. Although parenterally administered vaccines would induce a robust systemic immunity, they often provoke a weak mucosal immunity which may not be effective in preventing mucosal infection. In contrast, mucosal administration potentially offers the dual benefit of inducing potent mucosal and systemic immunity which would be more effective in offering protection against mucosal viral infection. There are however many challenges posed by the mucosal environment which impede successful mucosal vaccination. The development of an effective delivery system remains a major challenge to the successful exploitation of mucosal mRNA vaccination. Nonetheless, a number of delivery vehicles have been experimentally harnessed with different degrees of success in the mucosal delivery of mRNA vaccines. In this review, we provide a comprehensive overview of mRNA vaccines and summarise their application in the fight against emerging viral diseases with particular emphasis on COVID-19 mRNA platforms. Furthermore, we discuss the prospects and challenges of mucosal administration of mRNA-based vaccines, and we explore the existing experimental studies on mucosal mRNA vaccine delivery.
|
|
Scooped by
Gilbert C FAURE
July 28, 2022 4:49 AM
|
SARS-CoV-2 typically enters the body via the nose or mouth. Because of this, many think a nasal spray vaccine would do a better job of preventing COVID-19 infection and cutting transmission than an injection.
|
|
Scooped by
Gilbert C FAURE
July 26, 2022 5:24 AM
|
Lipid nanoparticles have limitations in inhaled drug delivery, including low pulmonary
bioavailability and unoptimized formulation. Lung-derived extracellular vesicles (Lung-Exos)
may be naturally equipped for drug delivery to the lung.
|
|
Scooped by
Gilbert C FAURE
July 26, 2022 4:29 AM
|
Dr Sudaxshina Murdan (UCL School of Pharmacy) and Dr Ruth Griffin (University of Nottingham) launch the Mucosal Vaccines Network.
|
|
Scooped by
Gilbert C FAURE
June 29, 2022 2:24 AM
|
Breastfeeding protects against mucosal infections in infants. The underlying mechanisms through which immunity develops in human milk following maternal infection with mucosal pathogens are not well understood. We simulated nasal mucosal influenza infection through live attenuated influenza vaccination (LAIV) and compared immune responses in milk to inactivated influenza vaccination (IIV). Transcriptomic analysis was performed on RNA extracted from human milk cells to evaluate differentially expressed genes and pathways on days 1 and 7 post-vaccination. Both LAIV and IIV vaccines induced influenza-specific IgA that persisted for at least 6 months. Regulation of type I interferon production, toll-like receptor, and pattern recognition receptor signaling pathways were highly upregulated in milk on day 1 following LAIV but not IIV at any time point. Upregulation of innate immunity in human milk may provide timely protection against mucosal infections until antigen-specific immunity develops in the human milk-fed infant.
|
|
Scooped by
Gilbert C FAURE
April 26, 2022 1:52 PM
|
Although SARS-CoV-2 infects the upper respiratory tract, we know little about the amount, type, and kinetics of antibodies (Ab) generated in the oral cavity in response to COVID-19 vaccination. We collected serum and saliva samples from participants receiving two doses of mRNA COVID-19 vaccines and measured the level of anti-SARS-CoV-2 Ab. We detected anti-Spike and anti-Receptor Binding Domain (RBD) IgG and IgA, as well as anti-Spike/RBD associated secretory component in the saliva of most participants after dose 1. Administration of a second dose of mRNA boosted the IgG but not the IgA response, with only 30% of participants remaining positive for IgA at this timepoint. At 6 months post-dose 2, these participants exhibited diminished anti-Spike/RBD IgG levels, although secretory component-associated anti-Spike Ab were more stable. Examining two prospective cohorts we found that participants who experienced breakthrough infections with SARS-CoV-2 variants had lower levels of vaccine-induced serum anti-Spike/RBD IgA at 2–4 weeks post-dose 2 compared to participants who did not experience an infection, whereas IgG levels were comparable between groups. These data suggest that COVID-19 vaccines that elicit a durable IgA response may have utility in preventing infection. Our study finds that a local secretory component-associated IgA response is induced by COVID-19 mRNA vaccination that persists in some, but not all participants. The serum and saliva IgA response modestly correlate at 2–4 weeks post-dose 2. Of note, levels of anti-Spike serum IgA (but not IgG) at this timepoint are lower in participants who subsequently become infected with SARS-CoV-2. As new surges of SARS-CoV-2 variants arise, developing COVID-19 booster shots that provoke high levels of IgA has the potential to reduce person-to-person transmission.
|
|
Scooped by
Gilbert C FAURE
March 5, 2022 9:58 AM
|
Background: Shigella spp. and enterotoxigenic Escherichia coli (ETEC) cause high morbidity and mortality worldwide, yet no licensed vaccines are available to prevent corresponding infections. A live attenuated non-invasive Shigella vaccine strain lacking LPS O-antigen and expressing the ETEC...
|
|
|
Scooped by
Gilbert C FAURE
November 17, 2022 4:53 AM
|
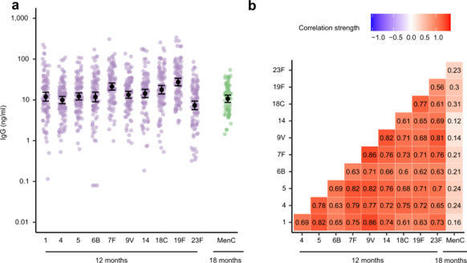
The gut microbiota in early life, when critical immune maturation takes place, may influence the immunogenicity of childhood vaccinations. Here we assess the association between mode of delivery, gut microbiota development in the first year of life, and mucosal antigen-specific antibody responses against pneumococcal vaccination in 101 infants at age 12 months and against meningococcal vaccination in 66 infants at age 18 months. Birth by vaginal delivery is associated with higher antibody responses against both vaccines. Relative abundances of vaginal birth-associated Bifidobacterium and Escherichia coli in the first weeks of life are positively associated with anti-pneumococcal antibody responses, and relative abundance of E. coli in the same period is also positively associated with anti-meningococcal antibody responses. In this study, we show that mode of delivery-induced microbiota profiles of the gut are associated with subsequent antibody responses to routine childhood vaccines. The establishment and composition of the host microbiota is known to impact the function of the host immune response. Here the authors show that mode of delivery may impact the intestinal microbiota composition from birth and modulate the response to routine childhood vaccines.
|
|
Scooped by
Gilbert C FAURE
November 5, 2022 7:04 AM
|
Mucosal Vaccines for SARS-CoV-2: Scientific Gaps and Opportunities. Online registration by Cvent
|
|
Scooped by
Gilbert C FAURE
October 12, 2022 6:10 AM
|
Systemic and mucosal humoral immune responses are crucial to fight respiratory viral infections in the current pandemic of COVID-19 caused by the SARS-CoV-2 virus. During SARS-CoV-2 infection, the dynamics of systemic and mucosal antibody infections are affected by patient characteristics, such as age, sex, disease severity, or prior immunity to other human coronaviruses. Patients suffering from severe disease develop higher levels of anti-SARS-CoV-2 antibodies in serum and mucosal tissues than those with mild disease, and these antibodies are detectable for up to a year after symptom onset. In hospitalized patients, the aberrant glycosylation of anti-SARS-CoV-2 antibodies enhances inflammation-associated antibody Fc-dependent effector functions, thereby contributing to COVID-19 pathophysiology. Current vaccines elicit robust humoral immune responses, principally in the blood. However, they are less effective against new viral variants, such as Delta and Omicron. This review provides an overview of current knowledge about the humoral immune response to SARS-CoV-2, with a particular focus on the protective and pathological role of humoral immunity in COVID-19 severity. We also discuss the humoral immune response elicited by COVID-19 vaccination and protection against emerging viral variants.
|
|
Scooped by
Gilbert C FAURE
October 5, 2022 12:26 PM
|
/PRNewswire/ -- Intravacc, a world leader in translational research and development of preventive and therapeutic vaccines, today announced that it has been...
|
|
Scooped by
Gilbert C FAURE
August 21, 2022 2:44 AM
|
A small spray in to the nose could protect people from COVID-19 and hamstring the pandemic, but the science still faces major challenges.
|
|
Scooped by
Gilbert C FAURE
August 18, 2022 5:07 AM
|
Longitudinal analysis of T cell, B cell, and antibody response to four different COVID-19 vaccines in humans, representing three different vaccine technologies, identifies different quantities and quality of CD4+ T cell, CD8+ T cell, and antibody response elicited by mRNA-1273, BNT162b2,...
|
|
Scooped by
Gilbert C FAURE
July 28, 2022 4:50 AM
|
Dozens of intranasally delivered vaccines targeting SARS-CoV-2 are in development. Could they pave the way for widespread nasal vaccination in the future?
|
|
Scooped by
Gilbert C FAURE
July 27, 2022 3:27 AM
|
The White House on Tuesday held a summit with vaccine makers and scientists as it pushed for "next generation" COVID-19 vaccines that offer broader and more durable protection against the virus.
|
|
Scooped by
Gilbert C FAURE
July 26, 2022 4:51 AM
|
When the vaccines were first introduced in December 2020, the virus they were designed against was altogether different from what it is today. SARS-CoV-2 had little substantive functional evolution from late 2019 until we saw the Alpha variant in the first months of 2021.
|
|
Scooped by
Gilbert C FAURE
July 22, 2022 3:59 AM
|
|
|
Scooped by
Gilbert C FAURE
May 23, 2022 11:03 AM
|
Mucosal vaccines offer the potential to trigger robust protective immune responses at the predominant sites of pathogen infection. In principle, the induction of adaptive immunity at mucosal sites, involving secretory antibody responses and tissue-resident T cells, has the capacity to prevent an infection from becoming established in the first place, rather than only curtailing infection and protecting against the development of disease symptoms. Although numerous effective mucosal vaccines are in use, the major advances seen with injectable vaccines (including adjuvanted subunit antigens, RNA and DNA vaccines) have not yet been translated into licensed mucosal vaccines, which currently comprise solely live attenuated and inactivated whole-cell preparations. The identification of safe and effective mucosal adjuvants allied to innovative antigen discovery and delivery strategies is key to advancing mucosal vaccines. Significant progress has been made in resolving the mechanisms that regulate innate and adaptive mucosal immunity and in understanding the crosstalk between mucosal sites, and this provides valuable pointers to inform mucosal adjuvant design. In particular, increased knowledge on mucosal antigen-presenting cells, innate lymphoid cell populations and resident memory cells at mucosal sites highlights attractive targets for vaccine design. Exploiting these insights will allow new vaccine technologies to be leveraged to facilitate rational mucosal vaccine design for pathogens including severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and for cancer. Here, Ed Lavelle and Ross Ward discuss the unique aspects of mucosal immunity that must be considered when developing effective mucosal vaccines. The authors highlight the key immune cell populations that are targeted by mucosal vaccination strategies and explain how innovative adjuvant and delivery approaches should lead to new vaccines for infectious diseases and cancers.

|
Suggested by
LIGHTING
March 26, 2022 5:34 AM
|
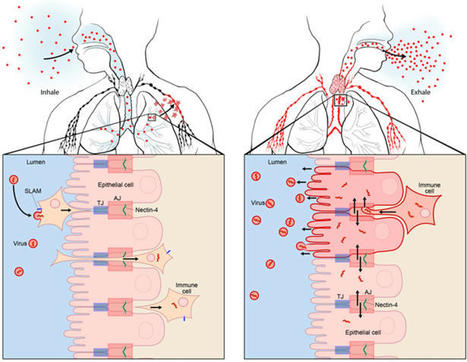
Measles VirusMeasles virus (MV) binds to cell surface receptors by way of its hemagglutinin (H) protein to initiate infection.From: xPharm: The Comprehensive Pharmacology Reference, 2007Related terms:Subacute Sclerosing PanencephalitisVaccine EfficacyCD46Monospecific AntibodyParamyxoviridaeImmunityWild TypeHuman Immunodeficiency VirusView all TopicsMeasles VirusR. Cattaneo, M. McChesney, in Encyclopedia of Virology (Third Edition), 2008Measles virus (MV), one of the most contagious viruses known, was recognized clinically by the rash and other signs from early historical times. Measles still causes more than 300 000 deaths each year, mostly due to secondary infections facilitated by MV-induced immunosuppression. Therefore the World Health Organization has targeted it for eradication, an endeavor facilitated by the availability of a live attenuated vaccine with an outstanding efficacy and safety record. MV is an enveloped negative-strand RNA virus, and the study of its biology has given detailed insights about how it utilizes host cell components to promote its replication and particle assembly. Moreover, the MV interactions with the cellular receptors and the mechanisms used to invade and inactivate the host immune system are now better understood. The knowledge gained from basic research is currently used to develop multivalent MV-based vaccines, and vectors for targeting and eliminating cancer cells.View chapterPurchase bookRead full chapterURL: https://www.sciencedirect.com/science/article/pii/B978012374410400443XDevelopmental Aspects of Pagetic OsteoclastsDeborah L. Galson, ... G. David Roodman, in Advances in Pathobiology and Management of Paget's Disease of Bone, 2016MVNP Downregulation of FoxO3 and Sirt1 also Increase IL-6MVNP also was demonstrated to increase NFκB activity by downregulating expression of Sirtuin 1 (Sirt1), a class III protein deacetylase that targets acetylated NFκB and negatively regulates its activity [31]. MVNP decreases Sirt1 by triggering increased phosphorylation of Forkhead-box class O3 (FoxO3), resulting in decreased FoxO3 protein stability and decreased transcription of its target gene Sirt1 in OCL precursors and NIH3T3 cells. Several protein kinases have been reported to downregulate FoxO3 stability through phosphorylation, including AKT, ERK1/2, IKKβ, and IKKε [50–52]. It’s not yet known which, if any, of these are triggered by MVNP to phosphorylate FoxO3. However, TBK1 overexpression in TRAP-TBK1 BMM was sufficient to decrease Sirt1 mRNA (unpublished data), suggesting that activated TBK1 may phosphorylate FoxO3. Wang et al. [31] showed that NIH3T3 cells stably transduced with MVNP (MVNP-NIH3T3) demonstrated higher IL-6 promoter luciferase reporter activity than NIH3T3 cells transduced with empty vector (EV-NIH3T3), and ectopic expression of Sirt1 significantly decreased both the basal and MVNP-stimulated IL-6 promoter activity. Further, resveratrol, a Sirt1 gene activator, suppressed the high level of IL-6 mRNA in MVNP-NIH3T3 cells. Significantly, resveratrol inhibited OCL differentiation of BMM from both wild-type and MVNP mice. Strikingly, at a resveratrol dose that had little effect on wild-type OCL differentiation, the enhanced MVNP OCL differentiation was suppressed to wild-type levels. Higher resveratrol doses then suppressed wild-type and MVNP OCL differentiation to similar levels. Hence, MVNP acts via two pathways to increase IL-6 expression (Fig. 4.2).View chapterPurchase bookRead full chapterURL: https://www.sciencedirect.com/science/article/pii/B978012805083500004XNeurovirologyJane E. Libbey, Robert S. Fujinami, in Handbook of Clinical Neurology, 2014Measles virusMeasles virus (Chapter 27) is a Morbillivirus, of the family Paramyxoviridae, and is an enveloped single-stranded negative-sense RNA virus (Nathanson and Gonzalez-Scarano, 2007). Measles virus is spread by the respiratory route (Nathanson and Gonzalez-Scarano, 2007). One neurologic complication of measles virus infection, thought to be related to dysregulation of the cellular immune responses, as it can occur in the absence of viral replication in the CNS, is postinfectious encephalomyelitis, which develops within weeks of infection (Johnson et al., 1983, 1984; Hirsch et al., 1984; Johnson, 1987). However, measles virus can infect and persist in neurons (Parra et al., 1999; Ramakrishna et al., 2002). In immunocompetent individuals a CD4 + T-cell population mediates elimination of measles virus from neurons, possibly through secretion of IFN-γ (Parra et al., 1999; Schneider-Schaulies et al., 2003). Failure of the immune response to eliminate measles virus-infected cells completely from the CNS can result in viral persistence (Schneider-Schaulies et al., 1999).Measles virus is capable of persisting in neurons as a defective variant that spreads from neuron to neuron directly, without passage through the extracellular environment (Schneider-Schaulies et al., 1999, 2003; Lawrence et al., 2000; Nathanson and Gonzalez-Scarano, 2007). In this way the virus avoids detection and elimination by circulating high titers of measles virus-specific neutralizing antibody. The persistence of measles virus in the CNS can result in a progressive fatal encephalitis called SSPE developing in immunocompetent individuals several months to years after infection and recovery from acute measles virus infection (Schneider-Schaulies et al., 1999, 2003; Nathanson and Gonzalez-Scarano, 2007). As such, SSPE is primarily a disease of childhood and young adulthood (Gilden, 1983; Wolinsky, 1990).As described above, oligoclonal IgG antibody bands, with measles virus-restricted specificities, are characteristically found in the serum and CSF of patients with SSPE (Mehta et al., 1994; Burgoon et al., 2006; Tschen et al., 2006). These oligoclonal bands are a hyperimmune response to measles virus antigens, which, despite their neutralizing activity, are unable to control the viral infection (Mehta et al., 1994). This inability of measles virus-specific antibodies to control measles virus infection in SSPE patients may result from antibody-induced antigenic modulation by the virus (Fujinami and Oldstone, 1980, 1983; Fujinami et al., 1984). The expression of some measles virus antigens within and on the surface of measles virus-infected cells is altered upon binding of measles virus-specific antibodies such that the synthesis, assembly, and maturation of the virions are altered. In this way the measles virus-infected cells may avoid detection and lysis and measles virus may persist. Thus, in the case of measles virus, the antibody arm of the adaptive immune response to the virus may play a role in the initiation of viral persistence (Fujinami and Oldstone, 1980, 1983; Fujinami et al., 1984).View chapterPurchase bookRead full chapterURL: https://www.sciencedirect.com/science/article/pii/B9780444534880000109MeaslesWilliam J. Moss, in Tropical Dermatology (Second Edition), 2017Natural History, Pathogenesis, and PathologyMeasles virus is primarily transmitted by respiratory droplets over short distances and, less commonly, by small-particle aerosols that remain suspended in the air for long periods of time. The time from infection to clinical disease is approximately 10 days to the onset of fever and 14 days to the onset of rash. Persons with measles are infectious for several days before and after the onset of rash, when levels of measles virus in blood and body fluids are highest and when the symptoms of cough, coryza, and sneezing are most severe. The host immune response at sites of virus replication is responsible for the signs and symptoms of measles.Host immune responses to measles virus are essential for viral clearance, clinical recovery, and the establishment of long-term immunity.7 The protective efficacy of antibodies to measles virus is illustrated by the immunity conferred to infants from passively acquired maternal antibodies and the protection of exposed, susceptible individuals following administration of anti-measles-virus immunoglobulin. The duration of protective immunity following wild-type measles virus infection is generally lifelong.8 The immune responses to measles virus infection are associated with depressed responses to unrelated (non-measles-virus) antigens lasting for several weeks to months. This state of immune suppression enhances susceptibility to secondary bacterial and viral infections causing pneumonia and diarrhea, and is likely responsible for much of measles-associated morbidity and mortality.9 Vitamin A deficiency is a recognized risk factor for severe measles. The vitamin is essential for the maintenance of normal epithelial tissues throughout the body; measles virus itself infects and damages these tissues.View chapterPurchase bookRead full chapterURL: https://www.sciencedirect.com/science/article/pii/B9780323296342000146Measles Virus (Rubeola)Anne A. Gershon, in Mandell, Douglas, and Bennett's Principles and Practice of Infectious Diseases (Eighth Edition), 2015Growth of Measles Virus in Tissue CultureMeasles virus was first successfully isolated in the laboratory by Enders and Peebles in 1954.18 The virus was initially propagated in primary human renal cells but later was cultivated in cultured simian kidney cells. Wild-type measles virus is rather difficult to propagate in vitro because it is slow growing, and only a limited number of types of cell cultures are permissive for the virus.16 Typically, cytopathic effects produced by measles virus in tissue cultures consist of stellate cells with increased refractility and, especially on passage, multinucleated syncytial giant cells containing intranuclear inclusions. In the absence of cytopathic effects, virus replication can also be detected by hemadsorption of rhesus monkey erythrocytes. Presumptive isolates of measles virus are identified by typing with monoclonal antibodies by using immunofluorescence or plaque reduction tests.3,19 Reverse-transcriptase polymerase chain reaction (RT-PCR) assays for measles virus are also available (see later).View chapterPurchase bookRead full chapterURL: https://www.sciencedirect.com/science/article/pii/B9781455748013001624MeaslesAlpay Azap, Filiz Pehlivanoglu, in Emerging Infectious Diseases, 20142 What is the Causative Agent? (Taxonomy and Description of the Agent)Measles virus (MV) is a member of the genus Morbillivirus of the family Paramyxoviridea.4 It is an enveloped, non-segmented, single-stranded, negative-sense RNA virus. Measles virions are seen as pleomorphic spheres with a diameter of 100–250 nm on electron microscopy. Measles virus encodes at least eight structural proteins, which have letter names: F, C, H, L, M, N, P, and V. Of these proteins, H (hemagglutinin) has a role in the attachment of the virus to host cells, and F (fusion) is involved in the spread of the virus from one cell to another.4Measles virus is closely related to the viruses causing diseases in animals such as canine and phocine distemper and rinderpest viruses. It is stated that MV adapted to humans when humans first began to domesticate animals in Mesopotamia around 3000 BC.1 Today, wild MV is pathogenic only for primates.View chapterPurchase bookRead full chapterURL: https://www.sciencedirect.com/science/article/pii/B9780124169753000261Infections that cause secondary immune deficiencyVincent Robert Bonagura, David Walter Rosenthal, in Stiehm's Immune Deficiencies (Second Edition), 2020Measles virus: temporary immunosuppressionMeasles virus (MV) continues to cause child morbidity and mortality worldwide, despite the availability and use of an effective live attenuated measles vaccine.10–14 Part of the reason why control of MV continues to be elusive is that it is highly contagious for susceptible individuals and there are difficulties with vaccine delivery. MV infection begins in the respiratory tract, spreads systemically in lymphoid, epithelial and endothelial cells, and ultimately infects multiple organs,15 causing a characteristic fever, rash, and conjunctivitis 10–14 days after respiratory infection (Fig. 49.3A and B). High fever, rhinorrhea and conjunctivitis typically precede the rash and the rash migrates from the head and neck to the hands and feet over 3–4 days. Many of these manifestations are caused by the immune response made to MV, and commonly this response clears MV in infected tissues and prevents re-infection for life (Fig. 49.3C and D). However, MV infection can cause several weeks of immune suppression after resolution in select individuals. This is the primary cause of measles-associated deaths: MV-induced secondary infection.16 Although vaccination against measles is very high in the United States, 92.7% of children aged 19–35 months were vaccinated in 2017,17 there are pockets of unvaccinated people who are susceptible to local outbreaks of measles. Since “herd immunity” is primarily effective when the vaccination rate is around 96%,18 vaccination levels below this level leave children and adults at risk for primary MV infection and secondary microbial infections.Measles was the first virus clearly identified to cause increased susceptibility to other microbial secondary infections. Most often, measles-associated deaths are caused by severe, overwhelming pneumonia and diarrhea.16 Suppression of delayed hypersensitivity has been identified in tuberculin-sensitized individuals many weeks after complete resolution of MV infection (Fig. 49.3C).19 Furthermore, several weeks after successful MV recovery, increased susceptibility to other infections has been reported, and T cell function and in vitro proliferation of T cells in response to mitogens has been shown to be markedly decreased (Fig. 49.4A and B).1,20,21 Immunosuppression occurs during a period of intense immune activation that occurs during the onset of the MV rash and anti-MV immune responses (Fig. 49.3C and D). Lymphopenia, skewing of Th2-like chemokine polarized responses, and suppression of lymphocyte proliferation have also been documented (Fig. 49.3D). MV infection causes decreases in T and B cells in the blood during the MV rash period.22–25 Altered trafficking and increased apoptosis of MV-infected and uninfected lymphocytes contribute to the development of lymphopenia.22,26–30 While lymphocyte numbers rapidly return to normal in the blood after the rash resolves, immunologic abnormalities persist.21,22,31,32 Immune suppression, Th2 cytokine polarization of CD4+ T cells, and Treg induction have been associated with indirect immunosuppression caused by MV infection.33,34 MV infection is also associated with suppression of IL-12 expression, lymphocyte CD30 expression, and IL-4, IL-10, and IL-13 expression after rash resolution.35–37 Reduction of IL-12 production reduces T cell expression of type I cytokines, particularly IFN-γ10,32 (Fig. 49.3D). It is possible that MV interacts with the complement regulatory molecule CD46 in polarizing Th2-like cytokine production, causing activation of signaling cascades that modify cell function, although this interaction is not firmly established.38,39 The MV-CD46 interaction may alter innate immunity by selectively downregulating receptor expression.40–46 This would increase susceptibility to complement-mediated lysis of MV-infected cells, and decrease antigen presenting cell production of IL-1247,48 and crosslinking of CD46 on T cells, leading to the induction of regulatory CD4+ T cells and enhanced IL-10 levels.49 These interactions would induce Th2-like polarization that would favor B cell maturation, provide lifelong MV antibody memory, and protect against MV re-infection. This polarization, however, would also depress APC activation and Th1-like responses to new pathogens.MV suppresses PBMC proliferation to mitogens after MV resolution, and this continues for several weeks (Fig. 49.4B).20,31 IL-2 supplementation can improve, but not fully restore, this responsiveness. This suggests that defective IL-2 expression is in part responsible for this proliferative defect.50 Cell cycle arrest in G1 after in vitro infection with MV is a recognized cause of hyporesponsiveness to mitogens.42,51–53 MV RNA can persist in PBMCs for months after MV resolution54,55 and may reduce mitogen proliferation, although this has not been established. The receptor used by wild-type MV to infect cells, CD150, is a dual function co-receptor for lymphocyte activation, and enhances IFN-γ expression.56–58 However, MV binding to CD150 can also downregulate receptor expression.59,60 T cell signaling through the MV glycoprotein complex of H and F1-F2 in the membranes of virions or MV-infected cells61–65 may also contribute to immunosuppression. This inhibitory signal prevents T cell S-phase entry for several days, and is independent of cell death, membrane fusion, soluble inhibitor production, or T cell infection.52,61,62,65–67 Thus, there is a delay in cell cycle progression and an accumulation of T cells in the G0/G1 phase.52,66,67 The mechanism by which H/F1-F2 suppresses mitogen-induced proliferation is unknown, but it is associated with MV-induced interference of T cell activation of phosphoinositide 3-kinase (PI3K) in T cells, or IL-2 receptor ligation.68 IL-2 added to MV-treated cells activates signal transducer and activator of transcription 3 (STAT3) but fails to activate Akt kinase, which is required for cell cycle progression.69 The modulatory effects of MV with glycoprotein complexes, and the downstream consequences of this interaction, have recently been summarized.10,68–70 While the relevance of these processes to the in vivo suppression of T cell lymphoproliferation remains to be identified. The combination of the established mechanisms leading to post-MV infection immunosuppression, and those that remain to be elucidated, cause, in select individuals, severe and on occasion, fatal secondary infection with other microbes.A key epidemiologic factor in measles-related deaths is vitamin A deficiency. In the developing world, the World Health Organization recommends vitamin A supplementation. Studies have demonstrated improved outcomes and suggest an effect on the mucosal barrier and also on improved T cell function though by an undefined mechanism.Read full chapterView PDFRead full chapterURL: https://www.sciencedirect.com/science/article/pii/B9780128167687000491MeaslesWilliam J. Moss, ... W. Harry Feinstone, in Vaccines for Biodefense and Emerging and Neglected Diseases, 2009Measles VirusMeasles virus is a spherical, nonsegmented, single-stranded, negative-sense RNA virus and a member of the Morbillivirus genus in the family of Paramyxoviridae. Other members of the Morbillivirus genus are rinderpest virus and canine distemper virus. Although RNA viruses have high mutation rates, measles virus is an antigenically monotypic virus, meaning that the surface proteins responsible for inducing protective immunity have retained their antigenic structure. The public health significance is that measles vaccines developed decades ago from a single measles virus strain remain protective worldwide. Measles virus is killed by ultraviolet light and heat. Attenuated measles vaccine viruses retain these characteristics, necessitating a cold chain for transportation and storage.The measles virus RNA genome consists of approximately 16,000 nucleotides and is enclosed in a lipid-containing envelope derived from the host cell. The genome encodes eight proteins, two of which (V and C) are nonstructural proteins and are transcribed from the phosphoprotein (P) gene. Of the six structural proteins, P, large protein (L), and nucleoprotein (N) form the nucleocapsid housing the viral RNA. The hemagglutinin protein (H), fusion protein (F), and matrix protein (M), together with lipids from the host cell membrane, form the viral envelope.The H protein interacts with F to mediate fusion of the viral envelope with the host cell membrane (Malvoisin and Wild, 1993). The primary function of the H protein is to bind to the host cellular receptors for measles virus. The two identified receptors are CD46 and CD150 (SLAM). CD46 is a complement regulatory molecule expressed on all nucleated cells in humans. SLAM, an acronym for signaling lymphocyte activation molecule, is expressed on activated T and B lymphocytes and antigen-presenting cells. The binding sites on H for these receptors overlap and strains of measles virus differ in the efficiency with which each is used. Wild-type measles virus binds to cells primarily through the cellular receptor SLAM whereas most vaccine strains bind to CD46; however, most measles virus strains can use both CD46 and SLAM as receptors during acute infection (Schneider et al., 2002). Additional, as yet unidentified receptors for measles virus exist on human endothelial and epithelial cells (Andres et al., 2003).Other measles virus proteins are involved in viral replication. The P protein regulates transcription, replication, and the efficiency with which the N assembles into nucleocapsids (Spehner et al., 1997). The M protein links ribonucleoproteins with envelope proteins during virion assembly. The functions of V and C proteins have not been clearly defined, but both appear to contribute to the virulence of measles virus by regulating transcription and sensitivity to the antiviral effects of interferon (IFN) α/β (Valsamakis et al., 1998; Patterson et al., 2000).Variability within the genome is sufficient to allow for molecular epidemiologic investigation. Genetic characterization of wild-type measles viruses is based on sequence analysis of the genes encoding the N and H proteins. One of the most variable regions of the measles virus genome is the 450-nucleotide sequence at the carboxy-terminal of the N protein, with up to 12% variability between wild-type viruses. The World Health Organization (WHO) recognizes 8 clades of measles virus (designated A through H) and 23 genotypes (World Health Organization, 2006). New genotypes likely will be identified with enhanced surveillance and molecular characterization. As measles control efforts intensify, molecular surveillance of circulating measles virus strains can be used to document interruption of measles virus transmission and to identify the source and transmission pathways of measles virus outbreaks (Rota and Bellini, 2003). Molecular epidemiologic tools also would be important in documenting deliberate bioterrorist introductions of wild-type or genetically modified measles virus strains.Read full chapterView PDFRead full chapterURL: https://www.sciencedirect.com/science/article/pii/B9780123694089000305Viruses as Infectious Agents: Human and Animal VirusesRoberto Cattaneo, Michael McChesney, in Encyclopedia of Virology (Fourth Edition), 2021Life CycleMeV begins its circuit through selected organs of the human body within alveolar macrophages and dendritic cells (DC), which express the primary receptor SLAM. These cells first transfer the infection through the epithelial barrier, and then spread it into lymphoid tissues (Fig. 4, left panel). SLAM was originally identified on activated B and T lymphocytes, but it is also expressed constitutively on immature thymocytes, memory T cells, and certain B cells. Subsets of other cell types, including monocytes and DC, also express SLAM. This cellular distribution overlaps with the susceptibility of different cell types to wild-type MeV infection. Another strong argument for the central role of SLAM in MeV tropism is the fact that three morbilliviruses (MeV, canine distemper virus, and rinderpest virus) enter cells via SLAM (human, canine, or bovine, respectively). Experiments in macaques revealed that the earliest target cells after intratracheal MeV inoculation are DCs and alveolar macrophages.More recently nectin-4, also called poliovirus receptor-like-4 (PVRL4), was shown to serve as the receptor for MeV spread in the upper airway epithelium (Fig. 4, right panel). This adherens junction protein interacts with H with five times higher affinity than SLAM. Nectin-4 sustains basolateral entry of MeV, and of all animal morbilliviruses examined, into upper airway epithelial cells, including those of the trachea.Contrary to the infections with other respiratory viruses, MeV enters the airway epithelium “en masse”: several days after host-to-host transmission, highly infected immune cells synchronously deliver large amounts of virus to the upper airways, for secondary amplification. Since the upper airways are the anatomical location most useful to support particle aerosolization, this two-phase mechanism of host invasion may account for the extremely contagious nature of MeV infection.The live attenuated MeV vaccine strain, Edmonston, can also use the regulator of complement activation membrane cofactor protein (MCP; CD46) as a receptor. The primary function of CD46 is to bind and promote inactivation of the C3b and C4b complement products, a process protecting human cells from lysis by autologous complement, a function that requires ubiquitous expression. Only tissue-culture adapted MeV interact with CD46, and indiscriminate cell entry through this protein correlates with MeV attenuation. A chimeric MeV expressing a vaccine strain H protein, which binds to CD46, is attenuated in the nonhuman primate, and productive infection is still confined to SLAM-expressing cells in vivo.View chapterPurchase bookRead full chapterURL: https://www.sciencedirect.com/science/article/pii/B9780128096338215069Rubeola VirusYvonne A. Maldonado, Avinash K. Shetty, in Principles and Practice of Pediatric Infectious Diseases (Fifth Edition), 2018Description of the PathogenMeasles virus (MV) is an enveloped, nonsegmented, negative-strand RNA virus belonging to the Morbillivirus genus in the Paramyxoviridae family. MV is closely related to rinderpest virus, a cattle pathogen that was eradicated in 2011.6 MV genome consists of 15,894 nucleotides and encodes eight proteins.7 The surface envelope glycoproteins and the hemagglutinin (H) and fusion (F) proteins are the main targets for development of neutralizing antibodies.8 The matrix (M) protein is important in virus assembly. The internal proteins, nucleoprotein (NP), polymerase phosphoprotein (P), and large protein (L), form the nucleocapsid. Two nonstructural proteins (C and V) regulate the cellular response to infection.7The receptor binding H protein interacts with F protein and mediates virus attachment and host cell entry. The F protein enhances cell-to-cell spread of the virus. Neutralizing antibodies are primarily directed against the H protein and confer lifelong immunity.8 There are three identified cellular entry receptors for MV. Human signaling lymphocyte activation molecule (i.e., hSLAM or CD150) is a common immune cell receptor for all MV strains, whereas membrane cofactor protein (i.e., MCP or CD46) is a receptor for vaccine strains.9 A third receptor, poliovirus receptor–like 4 (i.e., PVRL4 or Nectin 4) is expressed on the adherens junctions of epithelial cells.10Wild-type MV infects only primates. Although MV is a monotypic virus, minor genetic and antigenic variation occurs in some virus strains.11 These antigenic variations have not compromised long-lasting immunity. MV is inactivated by lipid solvents, heat, and light.View chapterPurchase bookRead full chapterURL: https://www.sciencedirect.com/science/article/pii/B9780323401814002279
|